

DNA DSB repair pathway choice: an orchestrated handover mechanism
- PMID: 24363387
- PMCID: PMC4064598
- DOI: 10.1259/bjr.20130685
DNA DSB repair pathway choice: an orchestrated handover mechanism
Abstract
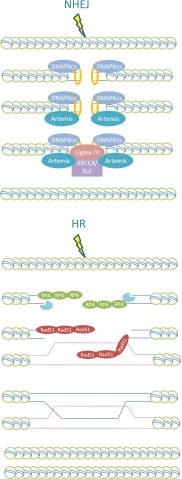
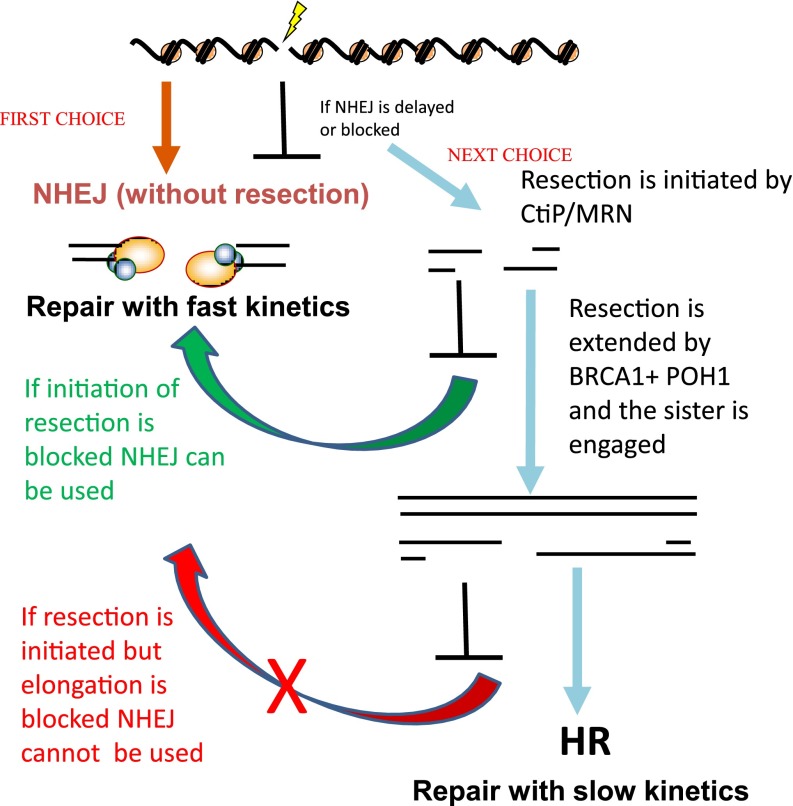
DNA double strand breaks (DSBs) are potential lethal lesions but can also lead to chromosome rearrangements, a step promoting carcinogenesis. DNA non-homologous end-joining (NHEJ) is the major DSB rejoining process and occurs in all cell cycle stages. Homologous recombination (HR) can additionally function to repair irradiation-induced two-ended DSBs in G2 phase. In mammalian cells, HR predominantly uses a sister chromatid as a template for DSB repair; thus HR functions only in late S/G2 phase. Here, we review current insight into the interplay between HR and NHEJ in G2 phase. We argue that NHEJ represents the first choice pathway, repairing approximately 80% of X-ray-induced DSBs with rapid kinetics. However, a subset of DSBs undergoes end resection and repair by HR. 53BP1 restricts resection, thereby promoting NHEJ. During the switch from NHEJ to HR, 53BP1 is repositioned to the periphery of enlarged irradiation-induced foci (IRIF) via a BRCA1-dependent process. K63-linked ubiquitin chains, which also form at IRIF, are also repositioned as well as receptor-associated protein 80 (RAP80), a ubiquitin binding protein. RAP80 repositioning requires POH1, a proteasome component. Thus, the interfacing barriers to HR, 53BP1 and RAP80 are relieved by POH1 and BRCA1, respectively. Removal of RAP80 from the IRIF core is required for loss of the ubiquitin chains and 53BP1, and for efficient replication protein A foci formation. We propose that NHEJ is used preferentially to HR because it is a compact process that does not necessitate extensive chromatin changes in the DSB vicinity.
Figures


References
-
- Lindahl T. Instability and decay of the primary structure of DNA. Nature 1993; 362: 709–14. - PubMed
-
- Friedberg EC. Out of the shadows and into the light: the emergence of DNA repair. Trends Biochem Sci 1995; 20: 381. - PubMed
-
- Sargentini NJ, Smith KC. Quantitation of the involvement of the recA, recB, recC, recF, recJ, recN, lexA, radA, radB, uvrD, and umuC genes in Escherichia coli. Radiat Res 1986; 107: 58–72. - PubMed
-
- Horii ZI, Clark AJ. Genetic analysis of the recF pathway to genetic recombination in Escherichia coli K-12: isolation and characterization of mutants. J Mol Biol 1973; 80: 327–44. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
