

Lineage specific antigenic differences in porcine torovirus hemagglutinin-esterase (PToV-HE) protein
- PMID: 24364900
- PMCID: PMC3878402
- DOI: 10.1186/1297-9716-44-126
Lineage specific antigenic differences in porcine torovirus hemagglutinin-esterase (PToV-HE) protein
Abstract
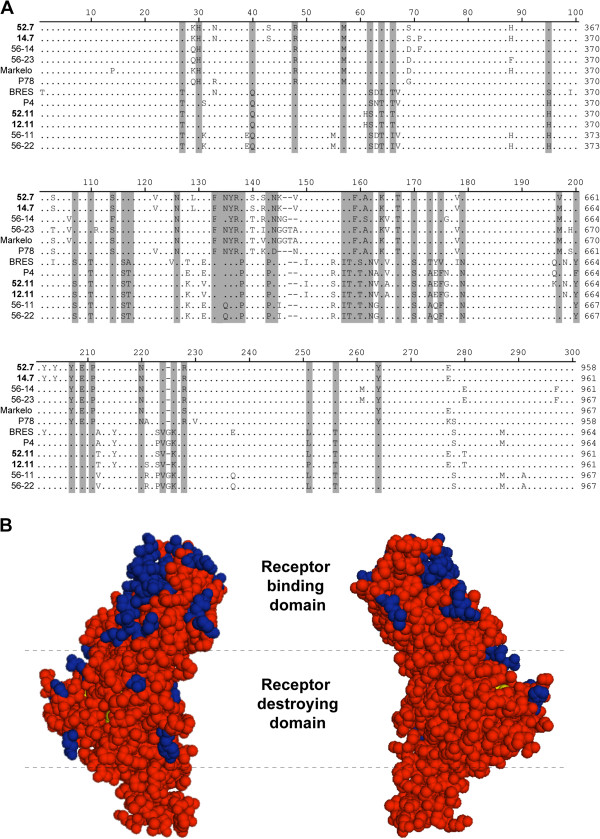
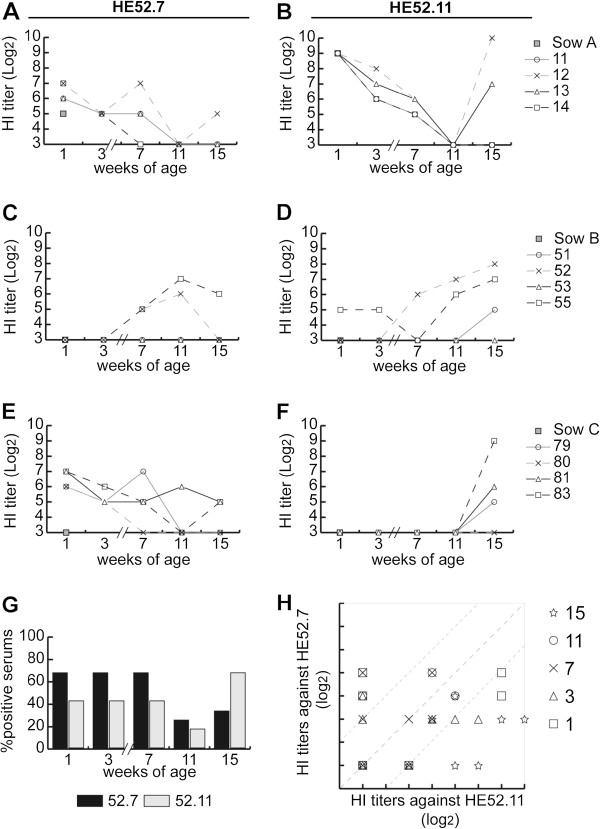
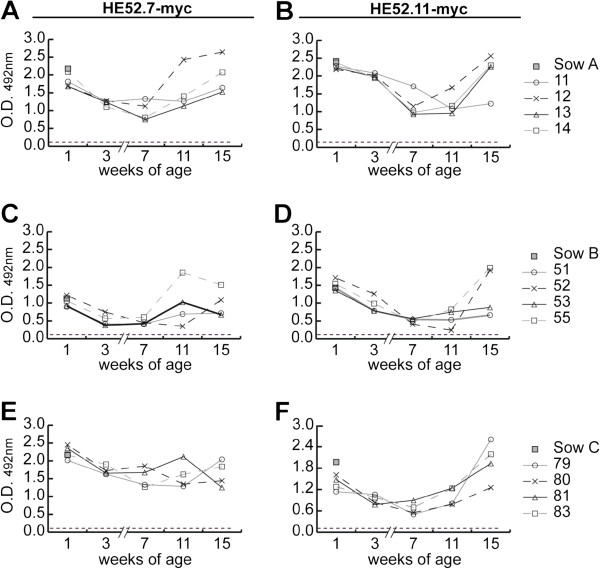
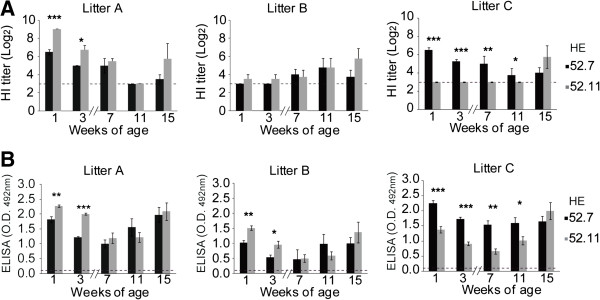
Hemagglutinin-esterases (HE) are viral envelope proteins present in some members from the toro-, corona- and orthomyxovirus families, all related with enteric and/or respiratory tract infections. HE proteins mediate reversible binding to sialic acid receptor determinants, very abundant glycan residues in the enteric and respiratory tracts. The role of the HE protein during the torovirus infection cycle remains unknown, although it is believed to be important in the natural infection process. The phylogenetic analysis of HE coding sequences from porcine torovirus (PToV) field strains revealed the existence of two distinct HE lineages. In a previous study, PToV virus strains with HE proteins from the two lineages were found coexisting in a pig herd, and they were even obtained from the same animal at two consecutive sampling time points. In this work, we report antigenic differences between the two HE lineages, and discuss the possible implications that the coexistence of viruses belonging to both lineages might have on the spread and sustainment of PToV infection in the farms.
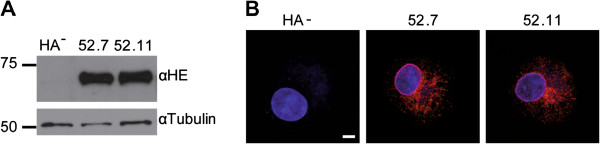
Figures
Similar articles
-
Evolution and homologous recombination of the hemagglutinin-esterase gene sequences from porcine torovirus.Virus Genes. 2013 Aug;47(1):66-74. doi: 10.1007/s11262-013-0926-y. Epub 2013 Jun 9. Virus Genes. 2013. PMID: 23749172 Free PMC article.
-
Molecular characterization of a new PToV strain. Evolutionary implications.Virus Res. 2009 Jul;143(1):33-43. doi: 10.1016/j.virusres.2009.02.019. Epub 2009 Mar 20. Virus Res. 2009. PMID: 19463719 Free PMC article.
-
Longitudinal serological and virological study on porcine torovirus (PToV) in piglets from Spanish farms.Vet Microbiol. 2010 Dec 15;146(3-4):260-8. doi: 10.1016/j.vetmic.2010.05.023. Epub 2010 May 16. Vet Microbiol. 2010. PMID: 20542392 Free PMC article.
-
[A review of porcine torovirus research: etiology and epidemiology].Bing Du Xue Bao. 2013 Nov;29(6):667-72. Bing Du Xue Bao. 2013. PMID: 24520775 Review. Chinese.
-
SARS-CoV-2 Evolutionary Adaptation toward Host Entry and Recognition of Receptor O-Acetyl Sialylation in Virus-Host Interaction.Int J Mol Sci. 2020 Jun 26;21(12):4549. doi: 10.3390/ijms21124549. Int J Mol Sci. 2020. PMID: 32604730 Free PMC article. Review.
Cited by
-
Recent Progress in Torovirus Molecular Biology.Viruses. 2021 Mar 8;13(3):435. doi: 10.3390/v13030435. Viruses. 2021. PMID: 33800523 Free PMC article. Review.
-
Activation of the autophagy pathway by Torovirus infection is irrelevant for virus replication.PLoS One. 2019 Jul 15;14(7):e0219428. doi: 10.1371/journal.pone.0219428. eCollection 2019. PLoS One. 2019. PMID: 31306441 Free PMC article.
-
Porcine Torovirus (PToV)-A Brief Review of Etiology, Diagnostic Assays and Current Epidemiology.Front Vet Sci. 2019 Apr 18;6:120. doi: 10.3389/fvets.2019.00120. eCollection 2019. Front Vet Sci. 2019. PMID: 31058174 Free PMC article. Review.
References
-
- Hoet A, Horzinek M. In: Encyclopedia of Virology. 3. Mahy BWJ, editor. Elsevier Academic Press; 2008. Torovirus; pp. 151–157.
-
- Waters V, Ford-Jones EL, Petric M, Fearon M, Corey P, Moineddein R. Etiology of community-acquired pediatric viral diarrhea: a prospective longitudinal study in hospitals, emergency departments, pediatric practices and child care centers during the winter rotavirus outbreak, 1997 to 1998. The pediatric rotavirus epidemiology study for immunization study group. Pediatr Infect Dis J. 2000;44:843–848. doi: 10.1097/00006454-200009000-00007. - DOI - PubMed
-
- Meng XJ. Emerging and re-emerging swine viruses. Transbound Emerg Dis. in press. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
