

Network mechanisms of grid cells
- PMID: 24366126
- PMCID: PMC3866436
- DOI: 10.1098/rstb.2012.0511
Network mechanisms of grid cells
Abstract
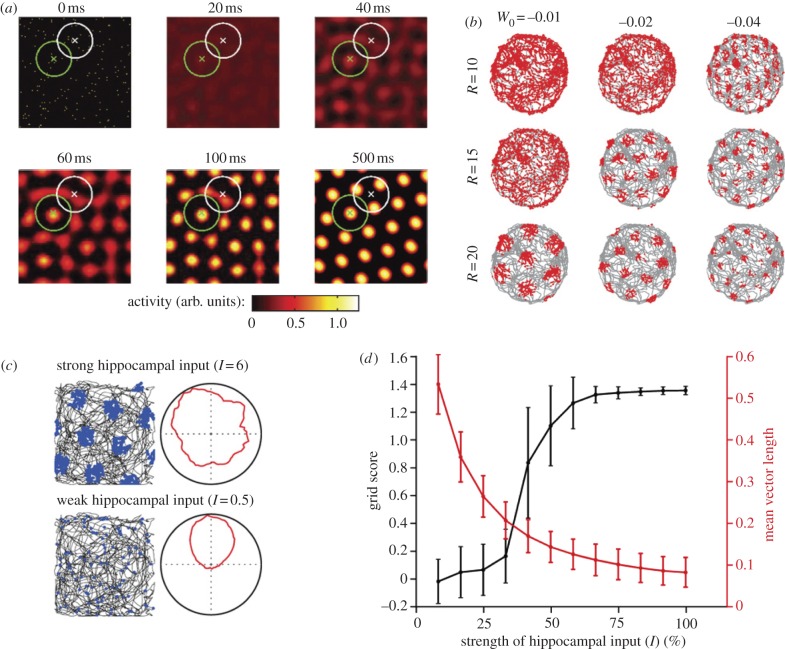
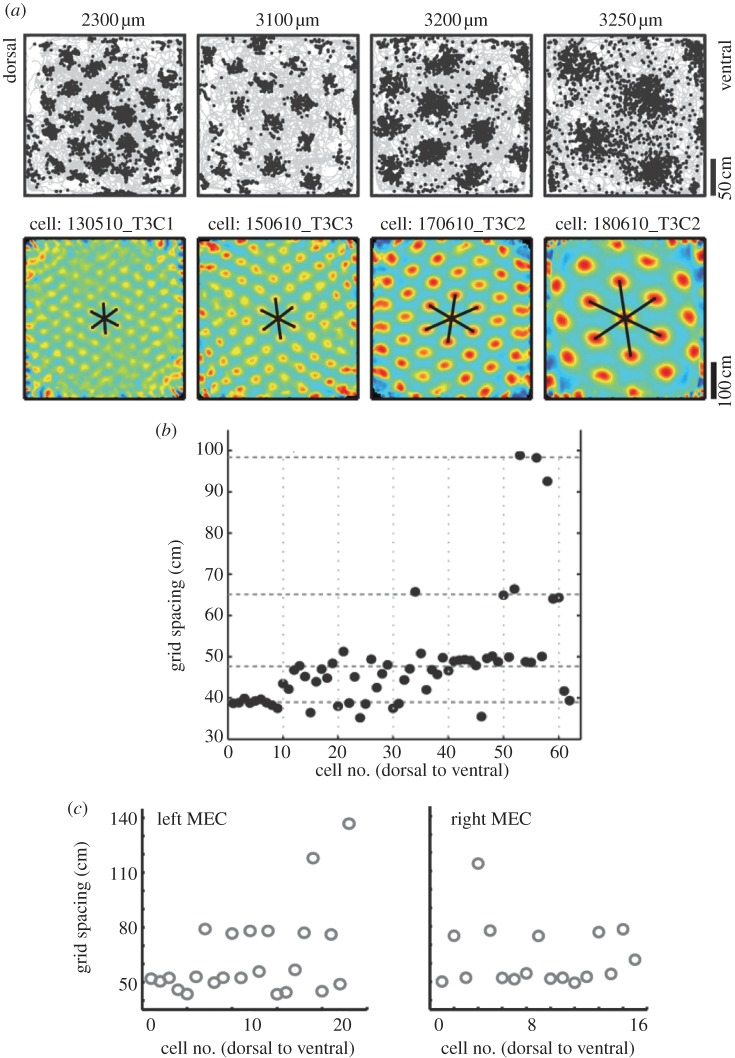
One of the major breakthroughs in neuroscience is the emerging understanding of how signals from the external environment are extracted and represented in the primary sensory cortices of the mammalian brain. The operational principles of the rest of the cortex, however, have essentially remained in the dark. The discovery of grid cells, and their functional organization, opens the door to some of the first insights into the workings of the association cortices, at a stage of neural processing where firing properties are shaped not primarily by the nature of incoming sensory signals but rather by internal self-organizing principles. Grid cells are place-modulated neurons whose firing locations define a periodic triangular array overlaid on the entire space available to a moving animal. The unclouded firing pattern of these cells is rare within the association cortices. In this paper, we shall review recent advances in our understanding of the mechanisms of grid-cell formation which suggest that the pattern originates by competitive network interactions, and we shall relate these ideas to new insights regarding the organization of grid cells into functionally segregated modules.
Keywords: attractor network; cortex; entorhinal cortex; grid cells; hippocampus; spatial representation.
Figures
References
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
