

Inhibition of cardiac pacemaker channel hHCN2 depends on intercalation of lipopolysaccharide into channel-containing membrane microdomains
- PMID: 24366264
- PMCID: PMC3961081
- DOI: 10.1113/jphysiol.2013.268540
Inhibition of cardiac pacemaker channel hHCN2 depends on intercalation of lipopolysaccharide into channel-containing membrane microdomains
Abstract
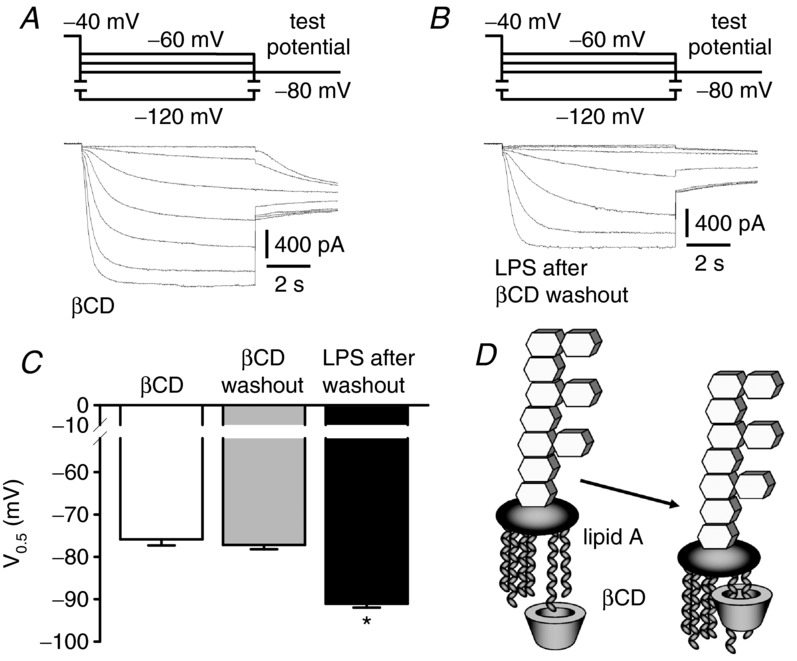
Depressed heart rate variability in severe inflammatory diseases can be partially explained by the lipopolysaccharide (LPS)-dependent modulation of cardiac pacemaker channels. Recently, we showed that LPS inhibits pacemaker current in sinoatrial node cells and in HEK293 cells expressing cloned pacemaker channels, respectively. The present study was designed to verify whether this inhibition involves LPS-dependent intracellular signalling and to identify structures of LPS responsible for pacemaker current modulation. We examined the effect of LPS on the activity of human hyperpolarization-activated cyclic nucleotide-gated channel 2 (hHCN2) stably expressed in HEK293 cells. In whole-cell recordings, bath application of LPS decreased pacemaker current (IhHCN2) amplitude. The same protocol had no effect on channel activity in cell-attached patch recordings, in which channels are protected from the LPS-containing bath solution. This demonstrates that LPS must interact directly with or close to the channel protein. After cleavage of LPS into lipid A and the polysaccharide chain, neither of them alone impaired IhHCN2, which suggests that modulation of channel activity critically depends on the integrity of the entire LPS molecule. We furthermore showed that β-cyclodextrin interfered with LPS-dependent channel modulation predominantly via scavenging of lipid A, thereby abrogating the capability of LPS to intercalate into target cell membranes. We conclude that LPS impairs IhHCN2 by a local mechanism that is restricted to the vicinity of the channels. Furthermore, intercalation of lipid A into target cell membranes is a prerequisite for the inhibition that is suggested to depend on the direct interaction of the LPS polysaccharide chain with cardiac pacemaker channels.
Figures
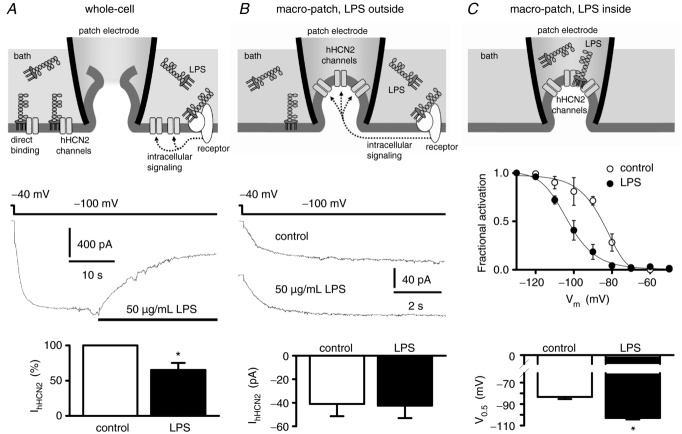
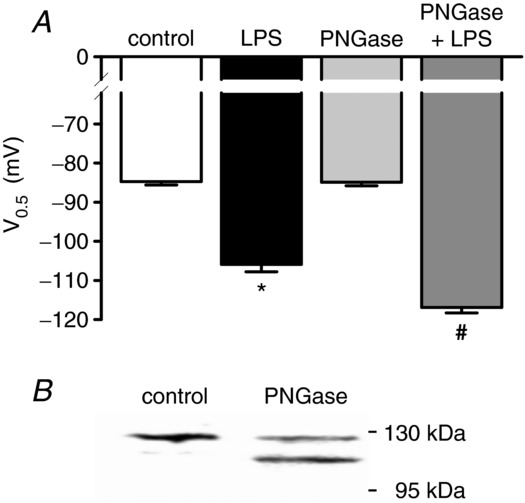
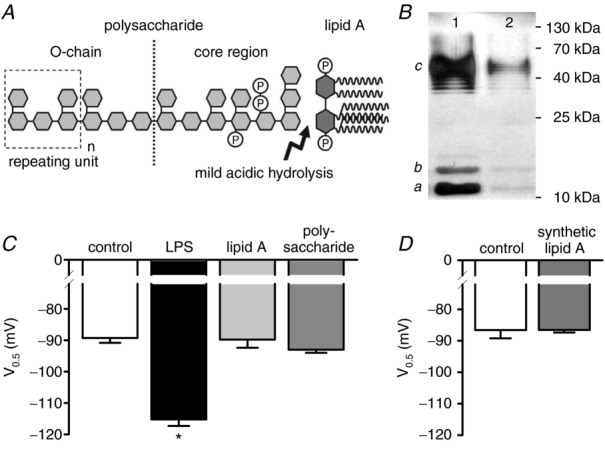
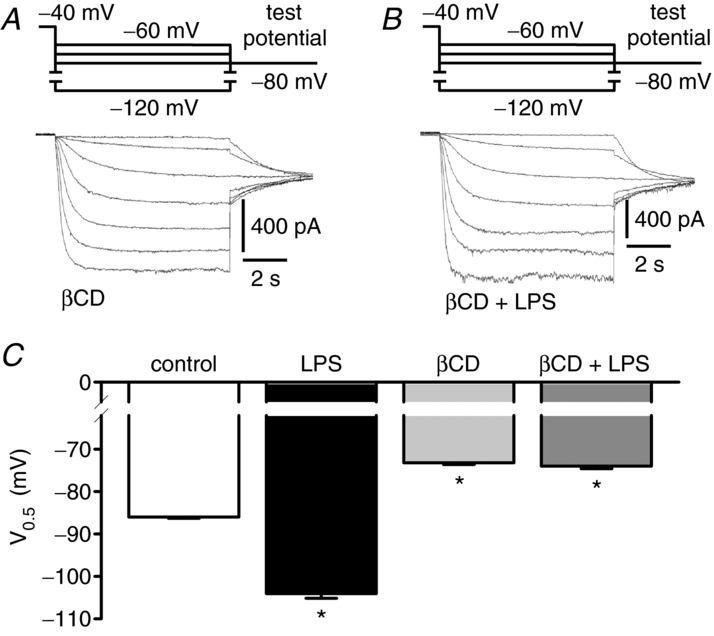
Comment in
-
The 'funny' side of sepsis.J Physiol. 2014 Mar 15;592(6):1171. doi: 10.1113/jphysiol.2014.271171. J Physiol. 2014. PMID: 24634015 Free PMC article. No abstract available.
References
-
- Alam JM, Yamazaki M. Spontaneous insertion of lipopolysaccharide into lipid membranes from aqueous solution. Chem Phys Lipids. 2011;164:166–174. - PubMed
-
- Allen JA, Halverson-Tamboli RA, Rasenick MM. Lipid raft microdomains and neurotransmitter signalling. Nat Rev Neurosci. 2007;8:128–140. - PubMed
-
- Barbuti A, Gravante B, Riolfo M, Milanesi R, Terragni B, DiFrancesco D. Localization of pacemaker channels in lipid rafts regulates channel kinetics. Circ Res. 2004;94:1325–1331. - PubMed
-
- Barbuti A, Scavone A, Mazzocchi N, Terragni B, Baruscotti M, DiFrancesco D. A caveolin-binding domain in the HCN4 channels mediates functional interaction with caveolin proteins. J Mol Cell Cardiol. 2012;53:187–195. - PubMed
-
- Baruscotti M, Bucchi A, Viscomi C, Mandelli G, Consalez G, Gnecchi-Rusconi T, Montano N, Casali KR, Micheloni S, Barbuti A, DiFrancesco D. Deep bradycardia and heart block caused by inducible cardiac-specific knock-out of the pacemaker channel gene Hcn4. Proc Natl Acad Sci U S A. 2011;108:1705–1710. - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources