

Evolution and functional cross-talk of protein post-translational modifications
- PMID: 24366814
- PMCID: PMC4019982
- DOI: 10.1002/msb.201304521
Evolution and functional cross-talk of protein post-translational modifications
Abstract
Protein post-translational modifications (PTMs) allow the cell to regulate protein activity and play a crucial role in the response to changes in external conditions or internal states. Advances in mass spectrometry now enable proteome wide characterization of PTMs and have revealed a broad functional role for a range of different types of modifications. Here we review advances in the study of the evolution and function of PTMs that were spurred by these technological improvements. We provide an overview of studies focusing on the origin and evolution of regulatory enzymes as well as the evolutionary dynamics of modification sites. Finally, we discuss different mechanisms of altering protein activity via post-translational regulation and progress made in the large-scale functional characterization of PTM function.
Figures
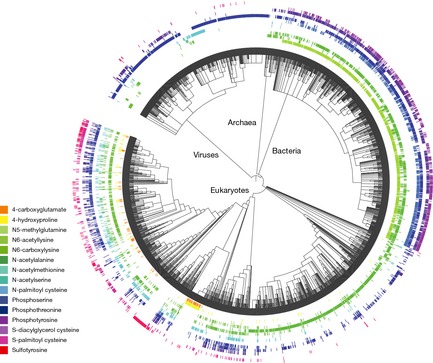
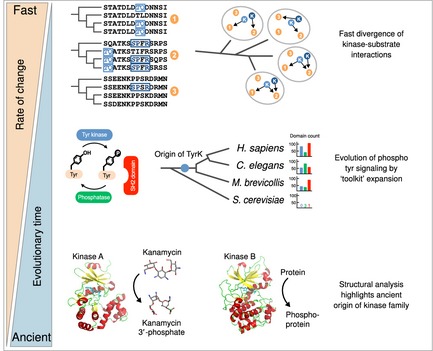
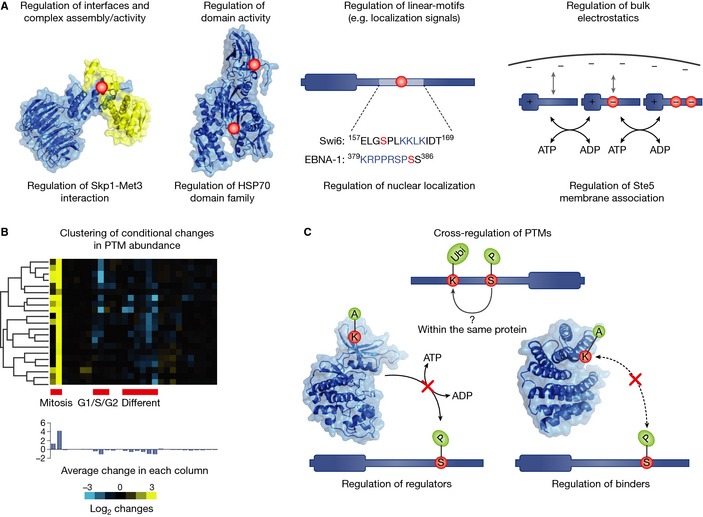
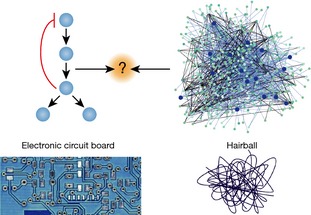
References
-
- Abu‐Qarn M, Eichler J, Sharon N (2008) Not just for Eukarya anymore: protein glycosylation in Bacteria and Archaea. Curr Opin Struct Biol 18: 544–550 - PubMed
-
- Alexander J, Lim D, Joughin BA, Hegemann B, Hutchins JRA, Ehrenberger T, Ivins F, Sessa F, Hudecz O, Nigg EA, Fry AM, Musacchio A, Stukenberg PT, Mechtler K, Peters J‐M, Smerdon SJ, Yaffe MB (2011) Spatial exclusivity combined with positive and negative selection of phosphorylation motifs is the basis for context‐dependent mitotic signaling. Sci Signal 4: ra42. - PMC - PubMed
-
- Anantharaman V, Iyer LM, Aravind L (2007) Comparative genomics of protists: new insights into the evolution of eukaryotic signal transduction and gene regulation. Annu Rev Microbiol 61: 453–475 - PubMed
-
- Bann JG, Bächinger HP (2000) Glycosylation/Hydroxylation‐induced stabilization of the collagen triple helix. 4‐trans‐hydroxyproline in the Xaa position can stabilize the triple helix. J Biol Chem 275: 24466–24469 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- P01AI090935/AI/NIAID NIH HHS/United States
- R01GM084448/GM/NIGMS NIH HHS/United States
- P01AI091575/AI/NIAID NIH HHS/United States
- R01 GM098101/GM/NIGMS NIH HHS/United States
- P50 GM082250/GM/NIGMS NIH HHS/United States
- R01 GM084448/GM/NIGMS NIH HHS/United States
- R01GM098101/GM/NIGMS NIH HHS/United States
- R01 GM084279/GM/NIGMS NIH HHS/United States
- P01 AI091575/AI/NIAID NIH HHS/United States
- P01 AI090935/AI/NIAID NIH HHS/United States
- R01GM084279/GM/NIGMS NIH HHS/United States
- P50GM081879/GM/NIGMS NIH HHS/United States
- P50 GM081879/GM/NIGMS NIH HHS/United States
- P50GM082250/GM/NIGMS NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
