

Trapping of naive lymphocytes triggers rapid growth and remodeling of the fibroblast network in reactive murine lymph nodes
- PMID: 24367096
- PMCID: PMC3890876
- DOI: 10.1073/pnas.1312585111
Trapping of naive lymphocytes triggers rapid growth and remodeling of the fibroblast network in reactive murine lymph nodes
Abstract
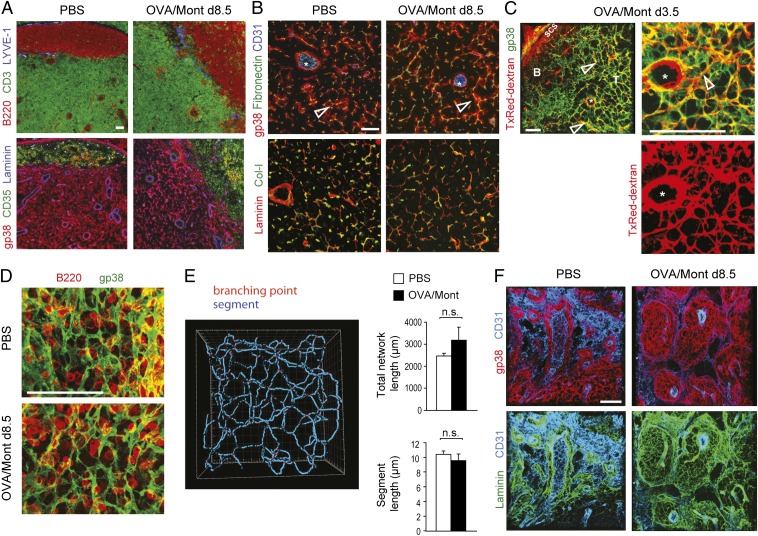
Adaptive immunity is initiated in T-cell zones of secondary lymphoid organs. These zones are organized in a rigid 3D network of fibroblastic reticular cells (FRCs) that are a rich cytokine source. In response to lymph-borne antigens, draining lymph nodes (LNs) expand several folds in size, but the fate and role of the FRC network during immune response is not fully understood. Here we show that T-cell responses are accompanied by the rapid activation and growth of FRCs, leading to an expanded but similarly organized network of T-zone FRCs that maintains its vital function for lymphocyte trafficking and survival. In addition, new FRC-rich environments were observed in the expanded medullary cords. FRCs are activated within hours after the onset of inflammation in the periphery. Surprisingly, FRC expansion depends mainly on trapping of naïve lymphocytes that is induced by both migratory and resident dendritic cells. Inflammatory signals are not required as homeostatic T-cell proliferation was sufficient to trigger FRC expansion. Activated lymphocytes are also dispensable for this process, but can enhance the later growth phase. Thus, this study documents the surprising plasticity as well as the complex regulation of FRC networks allowing the rapid LN hyperplasia that is critical for mounting efficient adaptive immunity.
Keywords: MyD88; fibroblasts; lymph node swelling; lymphotoxin; stromal cells.
Conflict of interest statement
The authors declare no conflict of interest.
Figures





References
-
- Turley SJ, Fletcher AL, Elpek KG. The stromal and haematopoietic antigen-presenting cells that reside in secondary lymphoid organs. Nat Rev Immunol. 2010;10(12):813–825. - PubMed
-
- Link A, et al. Fibroblastic reticular cells in lymph nodes regulate the homeostasis of naive T cells. Nat Immunol. 2007;8(11):1255–1265. - PubMed
-
- Angeli V, et al. B cell-driven lymphangiogenesis in inflamed lymph nodes enhances dendritic cell mobilization. Immunity. 2006;24(2):203–215. - PubMed
-
- Mohr E, et al. Dendritic cells and monocyte/macrophages that create the IL-6/APRIL-rich lymph node microenvironments where plasmablasts mature. J Immunol. 2009;182(4):2113–2123. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
