

Intestinal CD103+ dendritic cells are key players in the innate immune control of Cryptosporidium parvum infection in neonatal mice
- PMID: 24367259
- PMCID: PMC3868524
- DOI: 10.1371/journal.ppat.1003801
Intestinal CD103+ dendritic cells are key players in the innate immune control of Cryptosporidium parvum infection in neonatal mice
Abstract
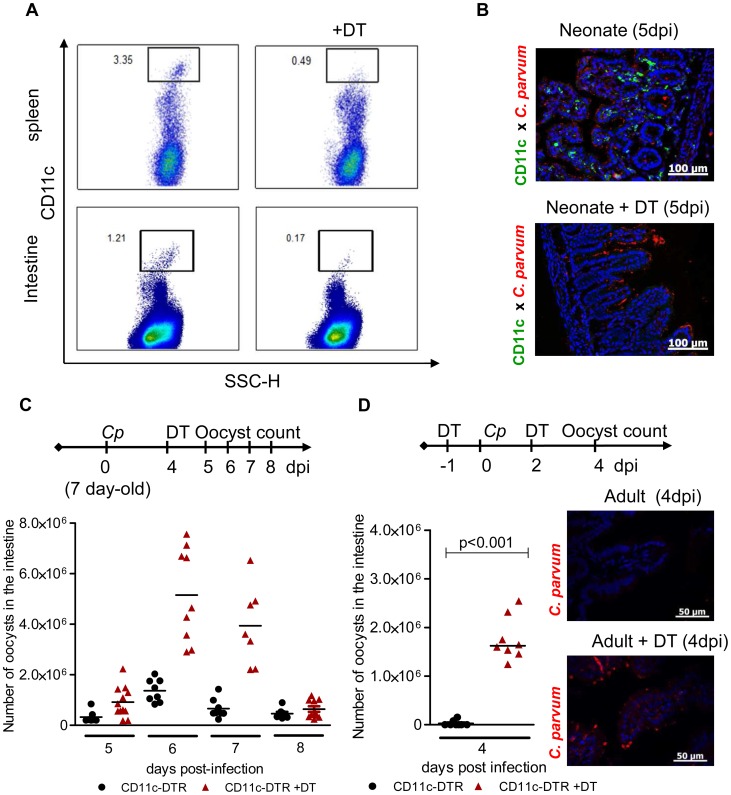
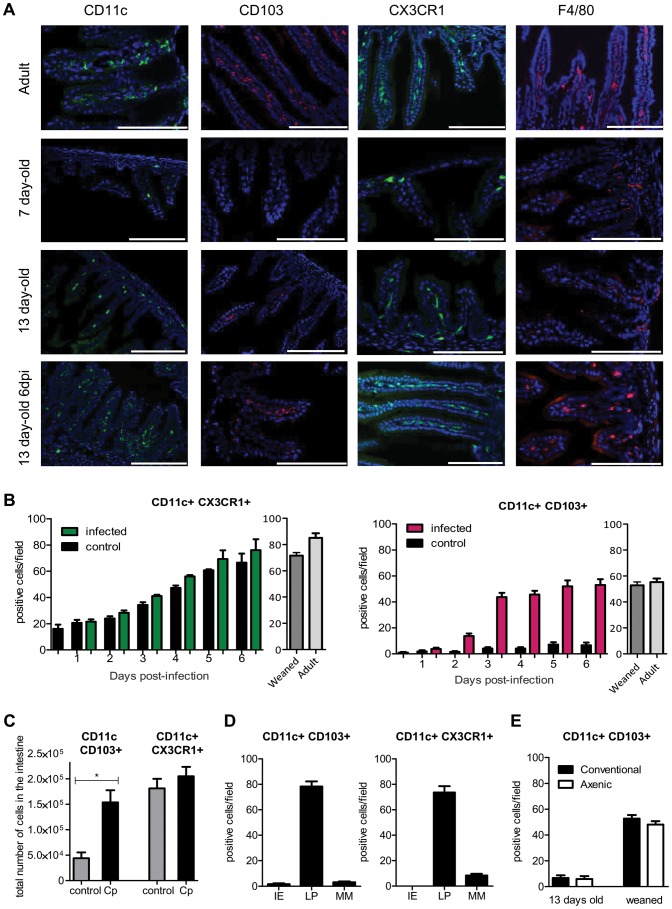
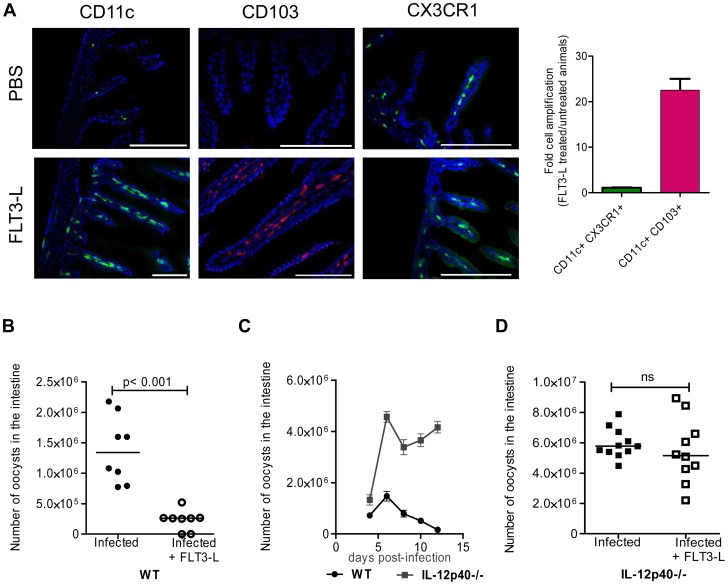
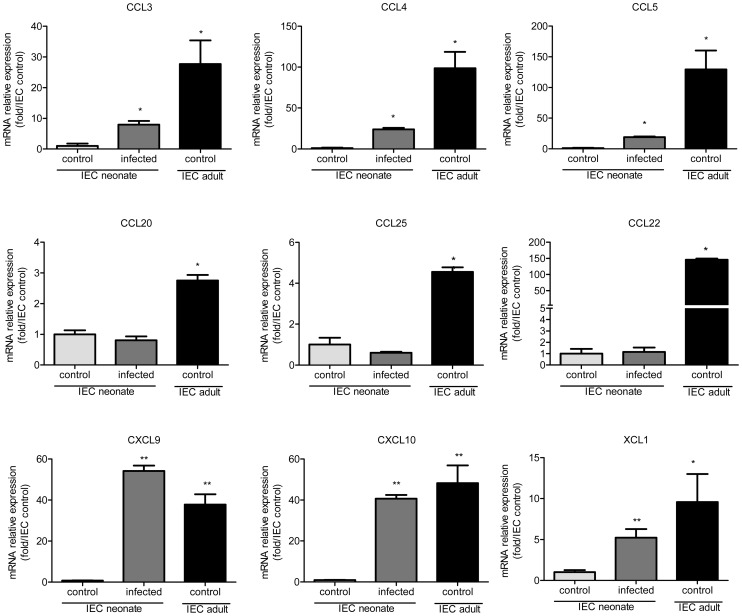
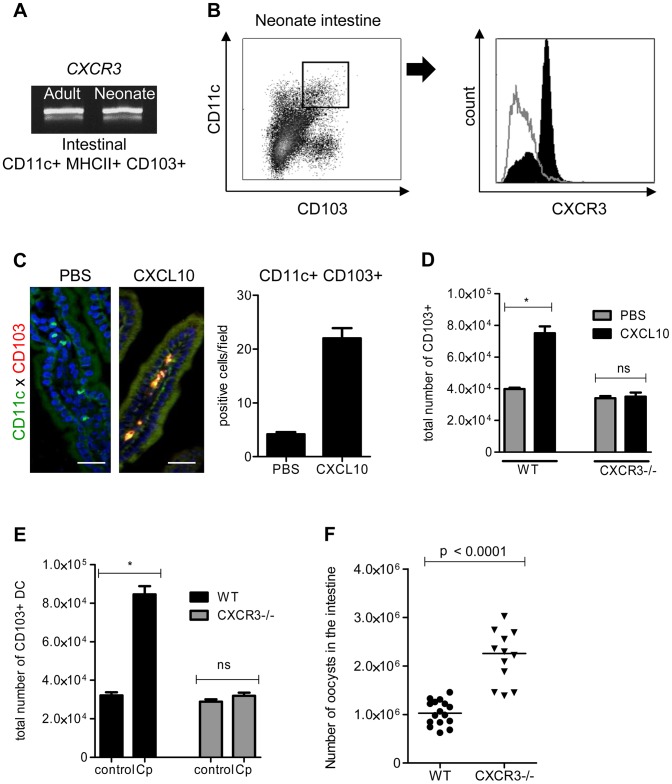
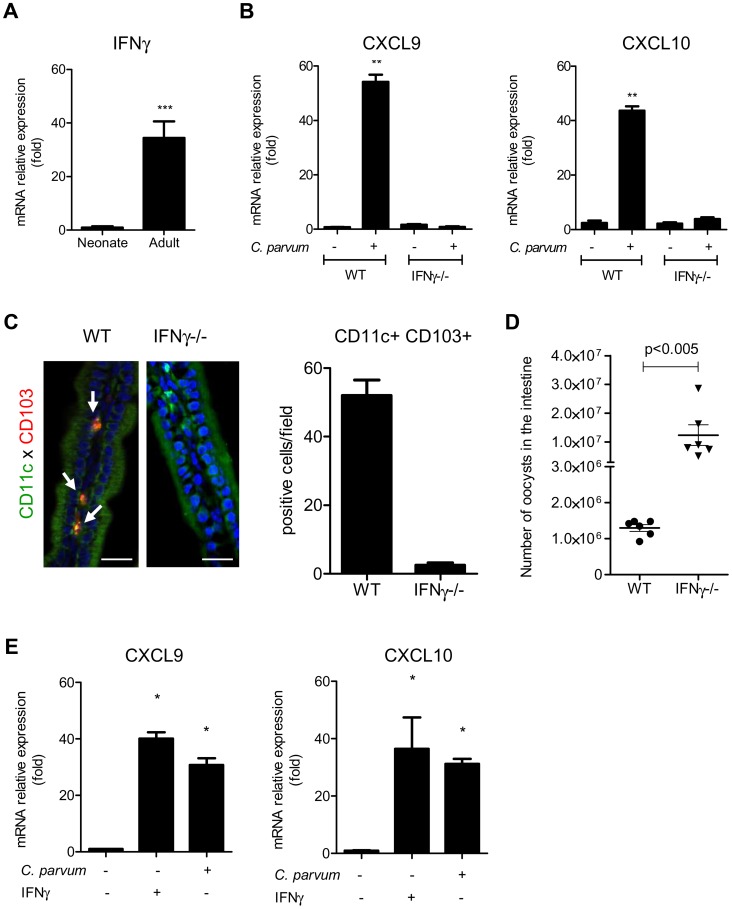
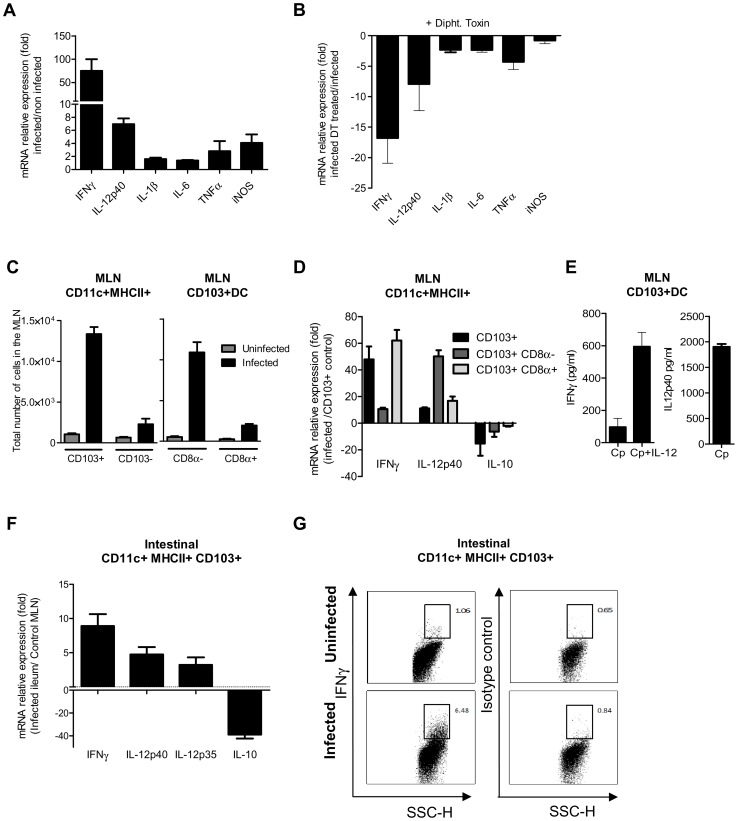
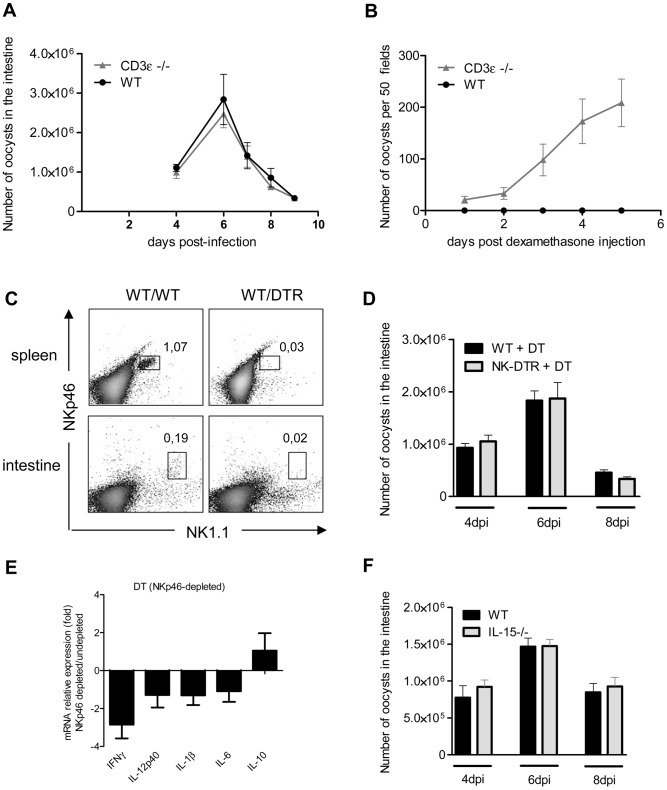
Cryptosporidium parvum is a zoonotic protozoan parasite found worldwide, that develops only in the gastrointestinal epithelium and causes profuse diarrhea. Using a mouse model of C. parvum infection, we demonstrated by conditional depletion of CD11c+ cells that these cells are essential for the control of the infection both in neonates and adults. Neonates are highly susceptible to C. parvum but the infection is self-limited, whereas adults are resistant unless immunocompromised. We investigated the contribution of DC to the age-dependent susceptibility to infection. We found that neonates presented a marked deficit in intestinal CD103+ DC during the first weeks of life, before weaning, due to weak production of chemokines by neonatal intestinal epithelial cells (IEC). Increasing the number of intestinal CD103+ DC in neonates by administering FLT3-L significantly reduced susceptibility to the infection. During infections in neonates, the clearance of the parasite was preceded by a rapid recruitment of CD103+ DC mediated by CXCR3-binding chemokines produced by IEC in response to IFNγ. In addition to this key role in CD103+ DC recruitment, IFNγ is known to inhibit intracellular parasite development. We demonstrated that during neonatal infection CD103+ DC produce IL-12 and IFNγ in the lamina propria and the draining lymph nodes. Thus, CD103+DC are key players in the innate immune control of C. parvum infection in the intestinal epithelium. The relative paucity of CD103+ DC in the neonatal intestine contributes to the high susceptibility to intestinal infection.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
-
- Chen XM, Keithly JS, Paya CV, LaRusso NF (2002) Cryptosporidiosis. N Engl J Med 346: 1723–1731. - PubMed
-
- Adkins B, Leclerc C, Marshall-Clarke S (2004) Neonatal adaptive immunity comes of age. Nat Rev Immunol 4: 553–564. - PubMed
-
- Renz H, Brandtzaeg P, Hornef M (2012) The impact of perinatal immune development on mucosal homeostasis and chronic inflammation. Nat Rev Immunol 12: 9–23. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
