

Biphasic euchromatin-to-heterochromatin transition on the KSHV genome following de novo infection
- PMID: 24367262
- PMCID: PMC3868514
- DOI: 10.1371/journal.ppat.1003813
Biphasic euchromatin-to-heterochromatin transition on the KSHV genome following de novo infection
Abstract
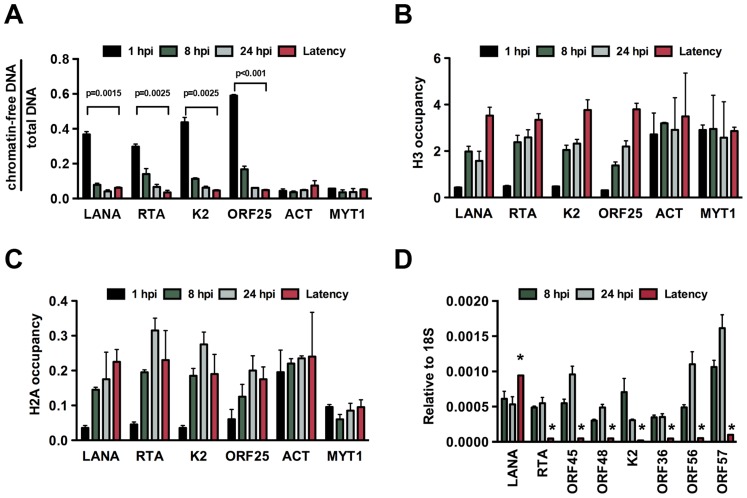
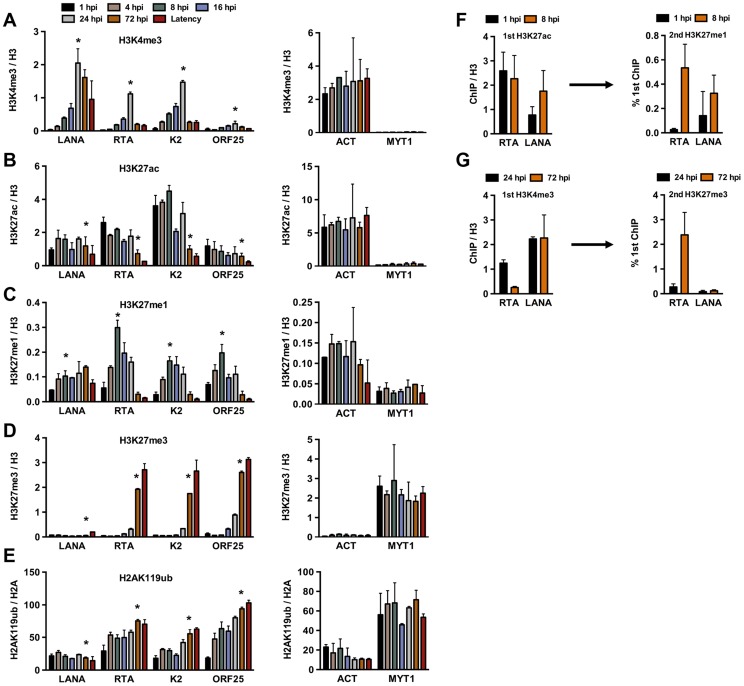
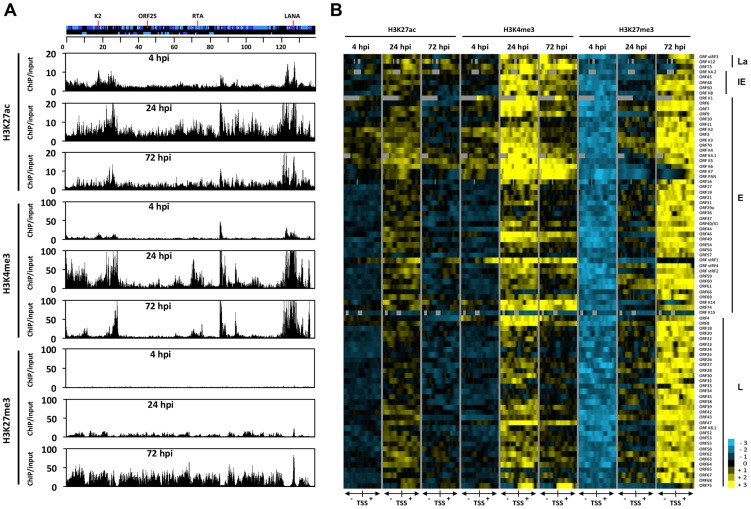
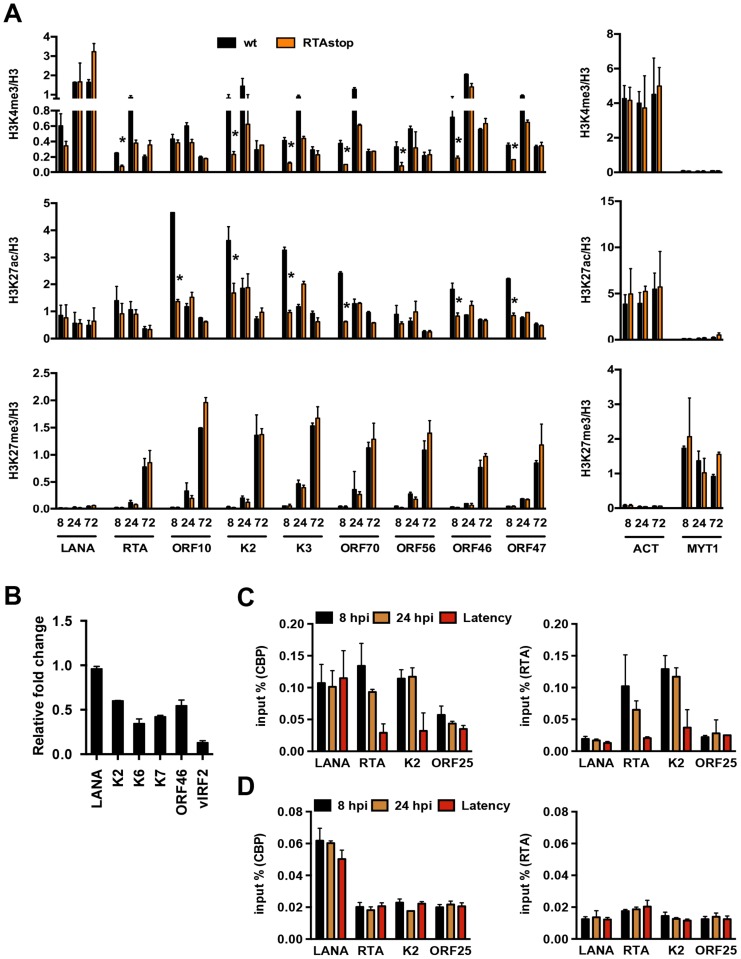
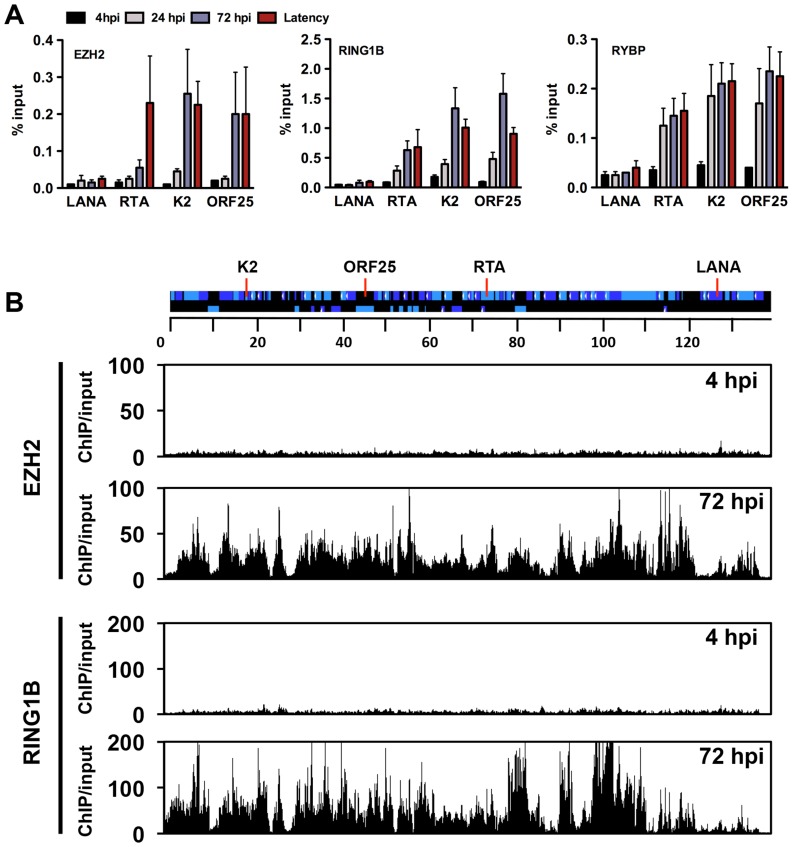
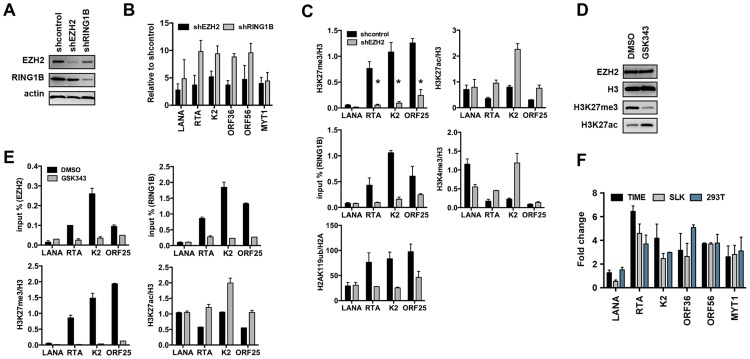
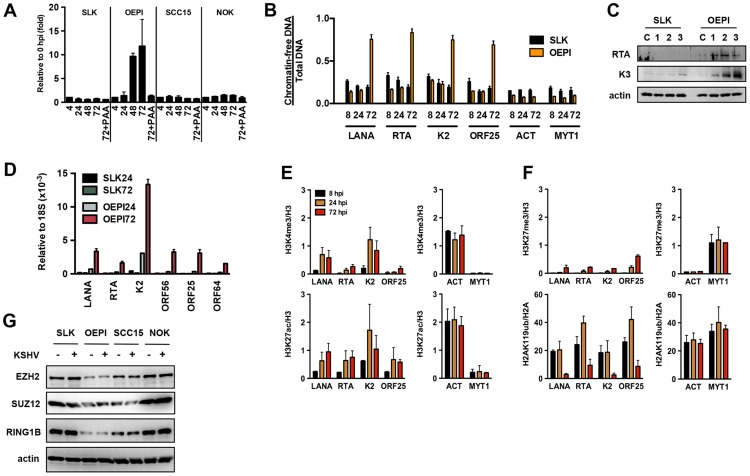
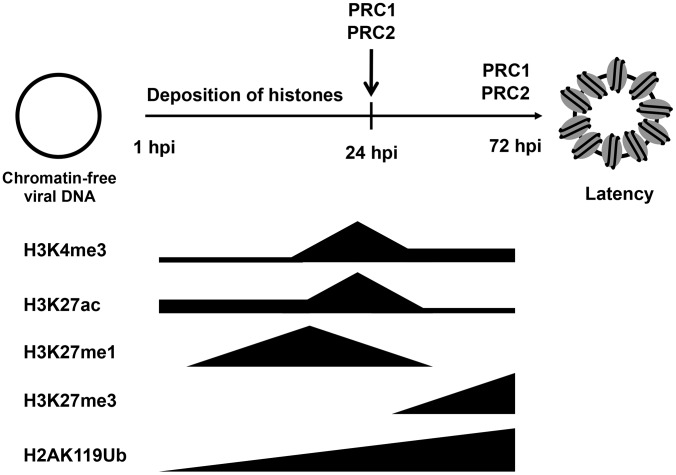
The establishment of latency is an essential step for the life-long persistent infection and pathogenesis of Kaposi's sarcoma-associated herpesvirus (KSHV). While the KSHV genome is chromatin-free in the virions, the viral DNA in latently infected cells has a chromatin structure with activating and repressive histone modifications that promote latent gene expression but suppress lytic gene expression. Here, we report a comprehensive epigenetic study of the recruitment of chromatin regulatory factors onto the KSHV genome during the pre-latency phase of KSHV infection. This demonstrates that the KSHV genome undergoes a biphasic chromatinization following de novo infection. Initially, a transcriptionally active chromatin (euchromatin), characterized by high levels of the H3K4me3 and acetylated H3K27 (H3K27ac) activating histone marks, was deposited on the viral episome and accompanied by the transient induction of a limited number of lytic genes. Interestingly, temporary expression of the RTA protein facilitated the increase of H3K4me3 and H3K27ac occupancy on the KSHV episome during de novo infection. Between 24-72 hours post-infection, as the levels of these activating histone marks declined on the KSHV genome, the levels of the repressive H3K27me3 and H2AK119ub histone marks increased concomitantly with the decline of lytic gene expression. Importantly, this transition to heterochromatin was dependent on both Polycomb Repressive Complex 1 and 2. In contrast, upon infection of human gingiva-derived epithelial cells, the KSHV genome underwent a transcription-active euchromatinization, resulting in efficient lytic gene expression. Our data demonstrate that the KSHV genome undergoes a temporally-ordered biphasic euchromatin-to-heterochromatin transition in endothelial cells, leading to latent infection, whereas KSHV preferentially adopts a transcriptionally active euchromatin in oral epithelial cells, resulting in lytic gene expression. Our results suggest that the differential epigenetic modification of the KSHV genome in distinct cell types is a potential determining factor for latent infection versus lytic replication of KSHV.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
-
- Cesarman E, Moore PS, Rao PH, Inghirami G, Knowles DM, et al. (1995) In vitro establishment and characterization of two acquired immunodeficiency syndrome-related lymphoma cell lines (BC-1 and BC-2) containing Kaposi's sarcoma-associated herpesvirus-like (KSHV) DNA sequences. Blood 86: 2708–2714. - PubMed
-
- Chang Y, Cesarman E, Pessin MS, Lee F, Culpepper J, et al. (1994) Identification of herpesvirus-like DNA sequences in AIDS-associated Kaposi's sarcoma. Science 266: 1865–1869. - PubMed
-
- Chang J, Renne R, Dittmer D, Ganem D (2000) Inflammatory cytokines and the reactivation of Kaposi's sarcoma-associated herpesvirus lytic replication. Virology 266: 17–25. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- P30 CA093373/CA/NCI NIH HHS/United States
- AI073099/AI/NIAID NIH HHS/United States
- CA31363/CA/NCI NIH HHS/United States
- R01 CA115284/CA/NCI NIH HHS/United States
- R01 AI073099/AI/NIAID NIH HHS/United States
- AI105809/AI/NIAID NIH HHS/United States
- CA180779/CA/NCI NIH HHS/United States
- R01 CA147791/CA/NCI NIH HHS/United States
- R01 CA082057/CA/NCI NIH HHS/United States
- R01 DE023926/DE/NIDCR NIH HHS/United States
- P01 CA180779/CA/NCI NIH HHS/United States
- DE023926/DE/NIDCR NIH HHS/United States
- CA082057/CA/NCI NIH HHS/United States
- CA115284/CA/NCI NIH HHS/United States
- R01 CA031363/CA/NCI NIH HHS/United States
- CA14779/CA/NCI NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
