

Functional analysis of ultra high information rates conveyed by rat vibrissal primary afferents
- PMID: 24367295
- PMCID: PMC3852094
- DOI: 10.3389/fncir.2013.00190
Functional analysis of ultra high information rates conveyed by rat vibrissal primary afferents
Abstract
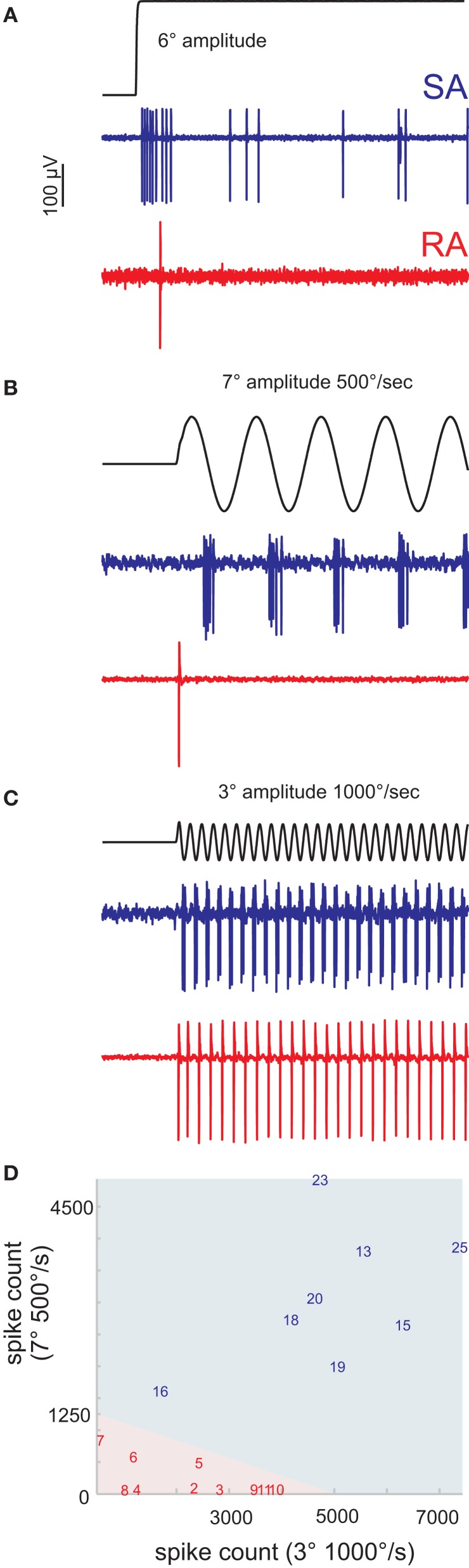
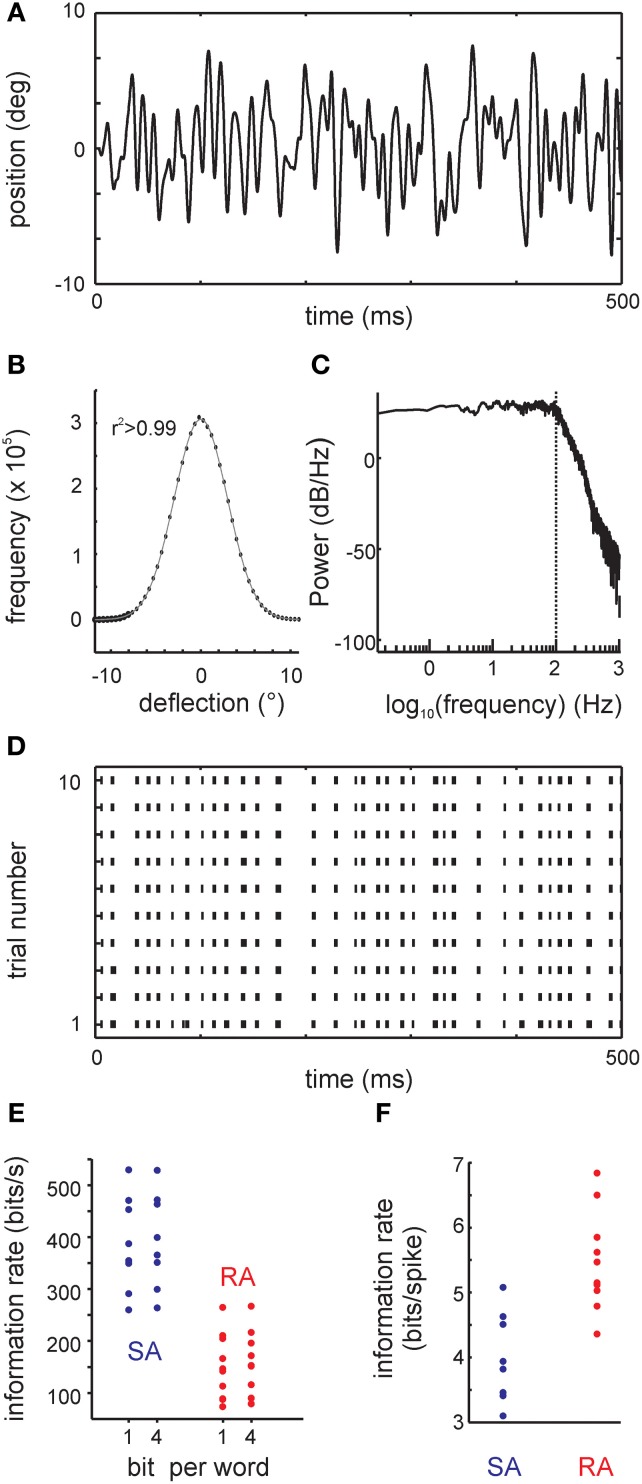
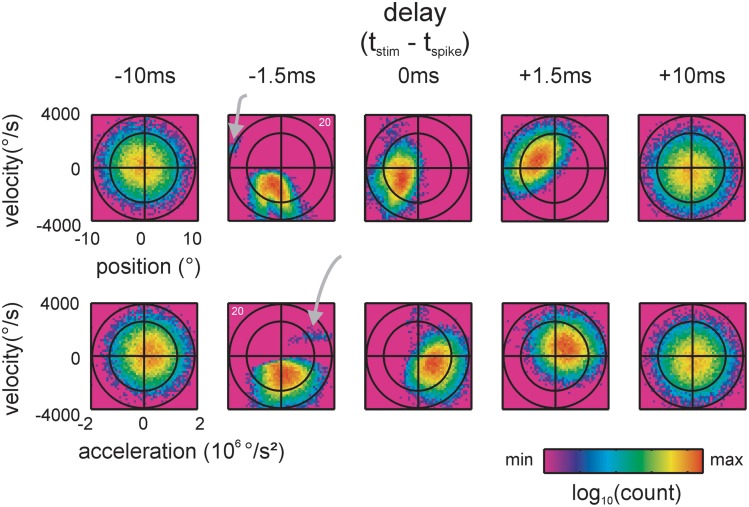
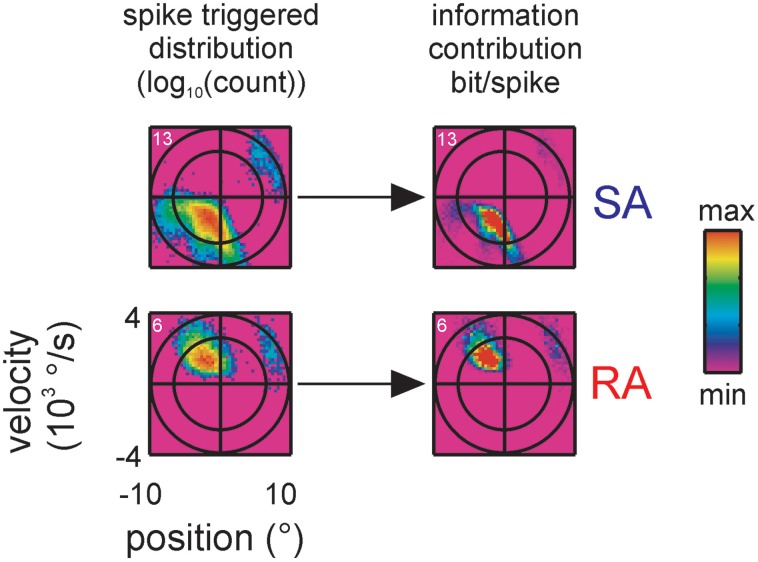
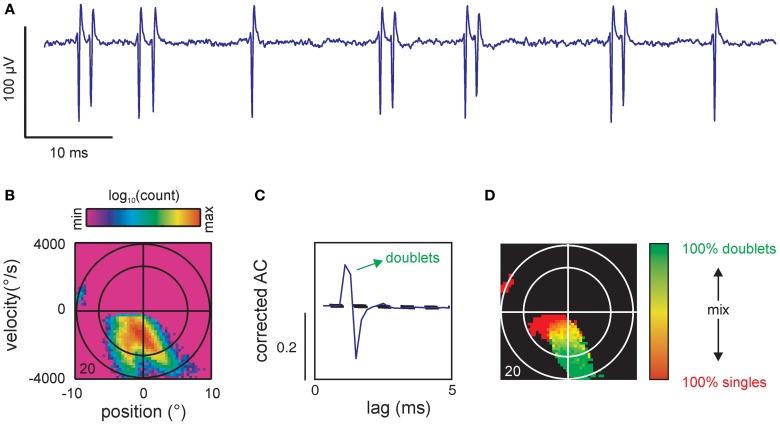
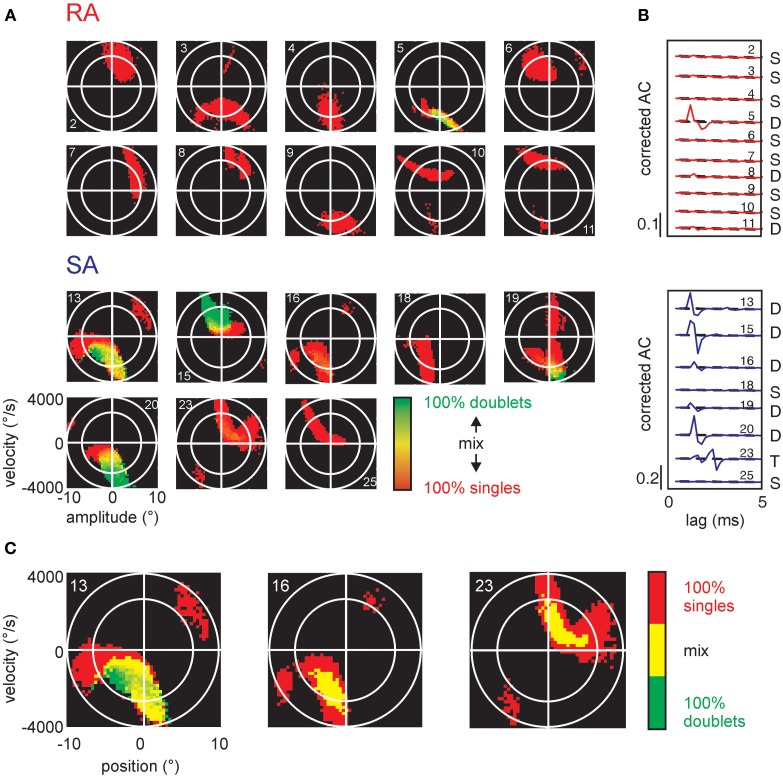
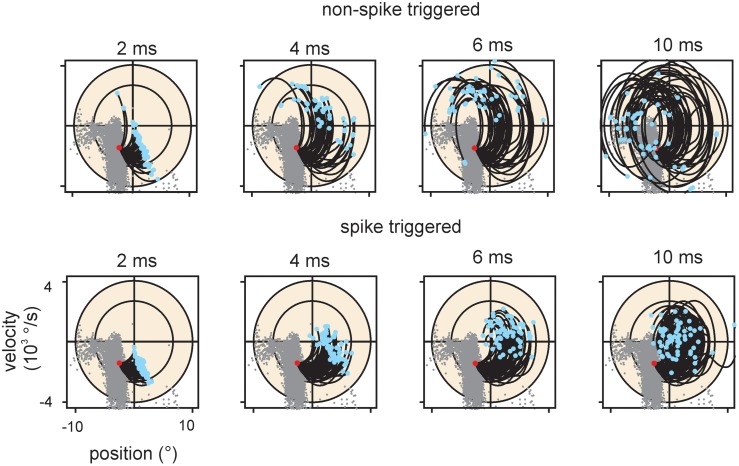
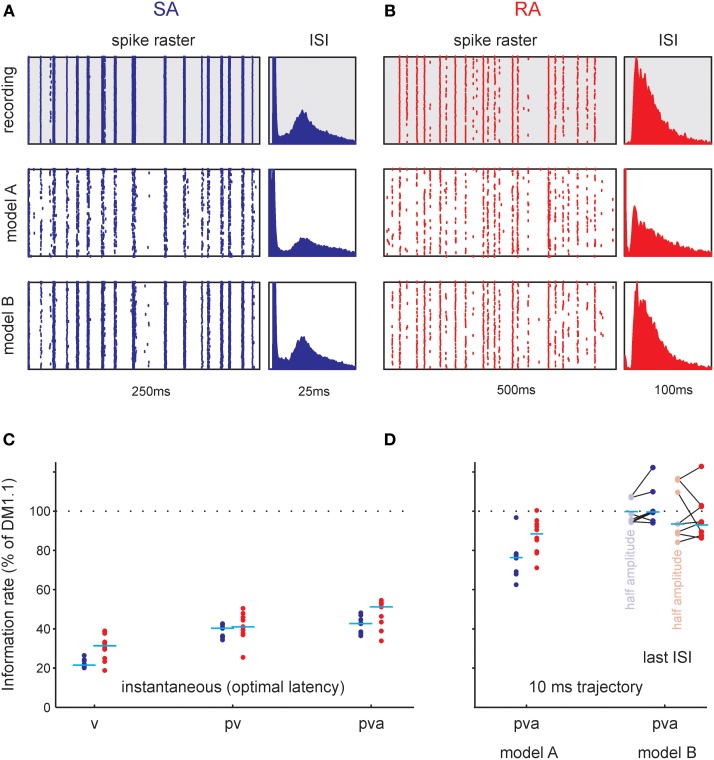
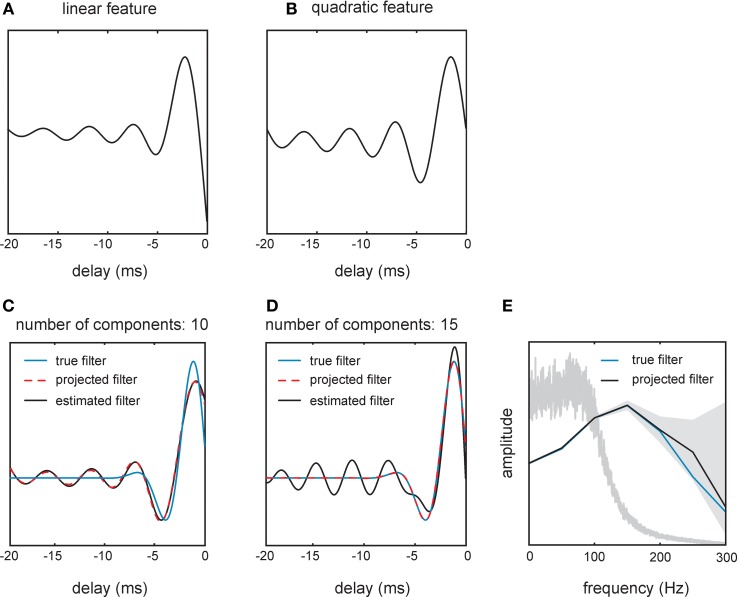
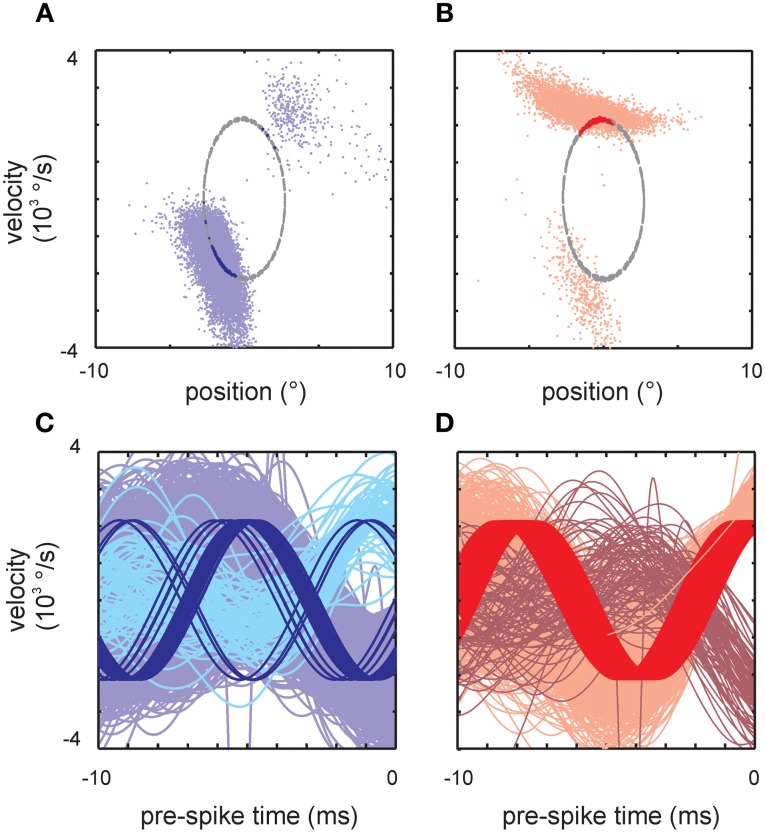
Sensory receptors determine the type and the quantity of information available for perception. Here, we quantified and characterized the information transferred by primary afferents in the rat whisker system using neural system identification. Quantification of "how much" information is conveyed by primary afferents, using the direct method (DM), a classical information theoretic tool, revealed that primary afferents transfer huge amounts of information (up to 529 bits/s). Information theoretic analysis of instantaneous spike-triggered kinematic stimulus features was used to gain functional insight on "what" is coded by primary afferents. Amongst the kinematic variables tested--position, velocity, and acceleration--primary afferent spikes encoded velocity best. The other two variables contributed to information transfer, but only if combined with velocity. We further revealed three additional characteristics that play a role in information transfer by primary afferents. Firstly, primary afferent spikes show preference for well separated multiple stimuli (i.e., well separated sets of combinations of the three instantaneous kinematic variables). Secondly, neurons are sensitive to short strips of the stimulus trajectory (up to 10 ms pre-spike time), and thirdly, they show spike patterns (precise doublet and triplet spiking). In order to deal with these complexities, we used a flexible probabilistic neuron model fitting mixtures of Gaussians to the spike triggered stimulus distributions, which quantitatively captured the contribution of the mentioned features and allowed us to achieve a full functional analysis of the total information rate indicated by the DM. We found that instantaneous position, velocity, and acceleration explained about 50% of the total information rate. Adding a 10 ms pre-spike interval of stimulus trajectory achieved 80-90%. The final 10-20% were found to be due to non-linear coding by spike bursts.
Keywords: information theory; primary afferents; rat; spike-triggered mixture model; tactile coding; vibrissae; whisker.
Figures


References
-
- Bishop C. M. (2006). Pattern Recognition and Machine Learning. New York, NY: Springer
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources