

Mouse Grueneberg ganglion neurons share molecular and functional features with C. elegans amphid neurons
- PMID: 24367309
- PMCID: PMC3856774
- DOI: 10.3389/fnbeh.2013.00193
Mouse Grueneberg ganglion neurons share molecular and functional features with C. elegans amphid neurons
Abstract
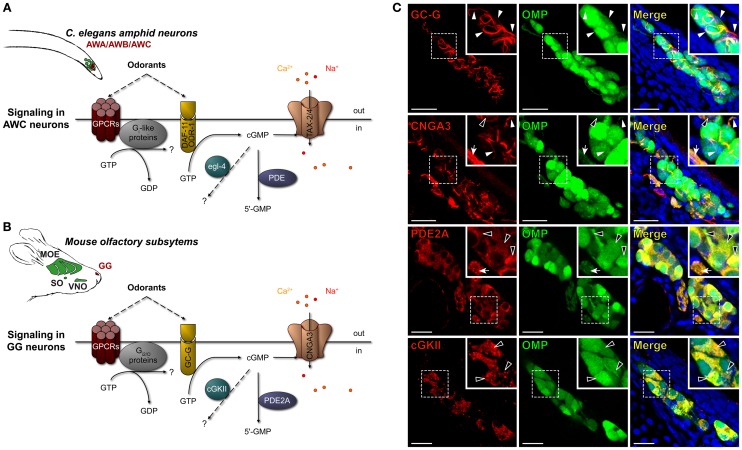
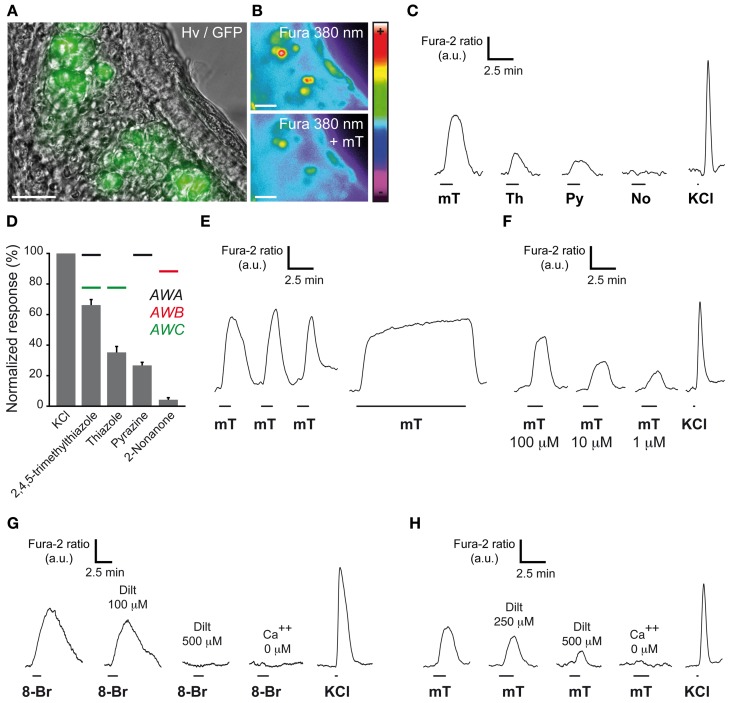
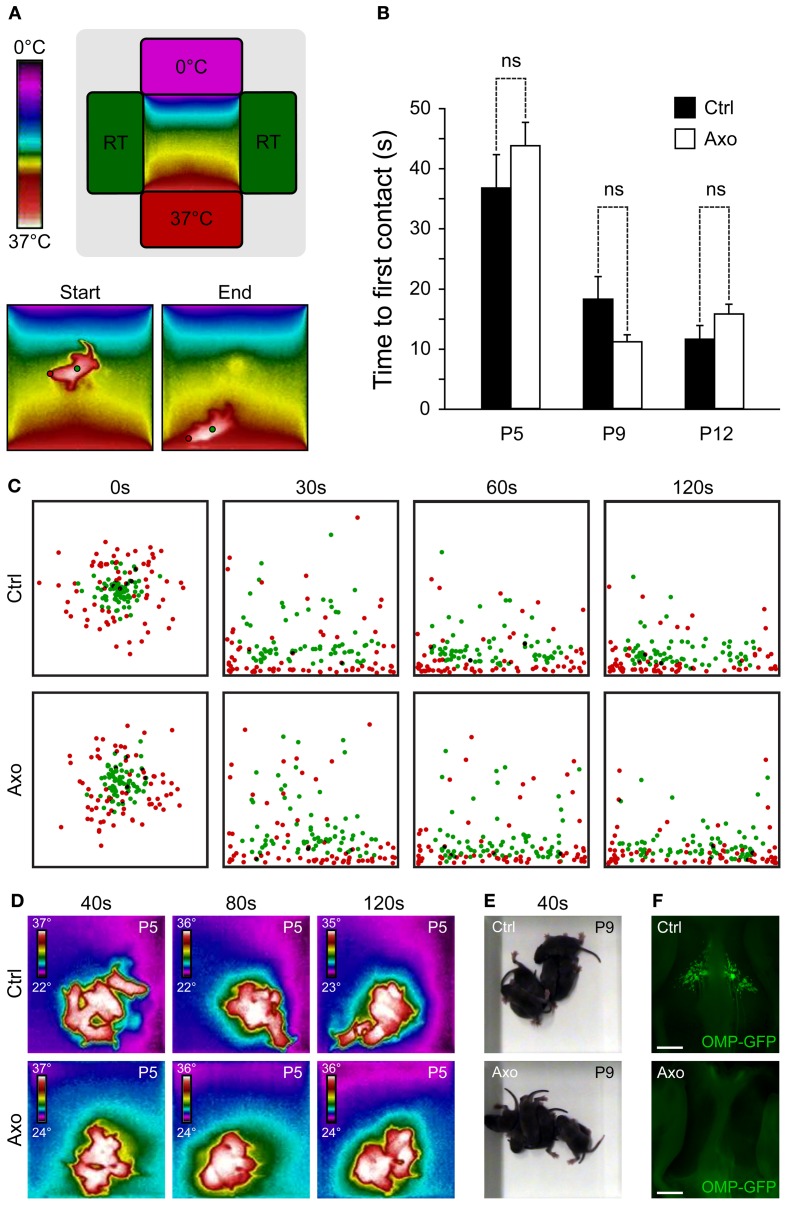
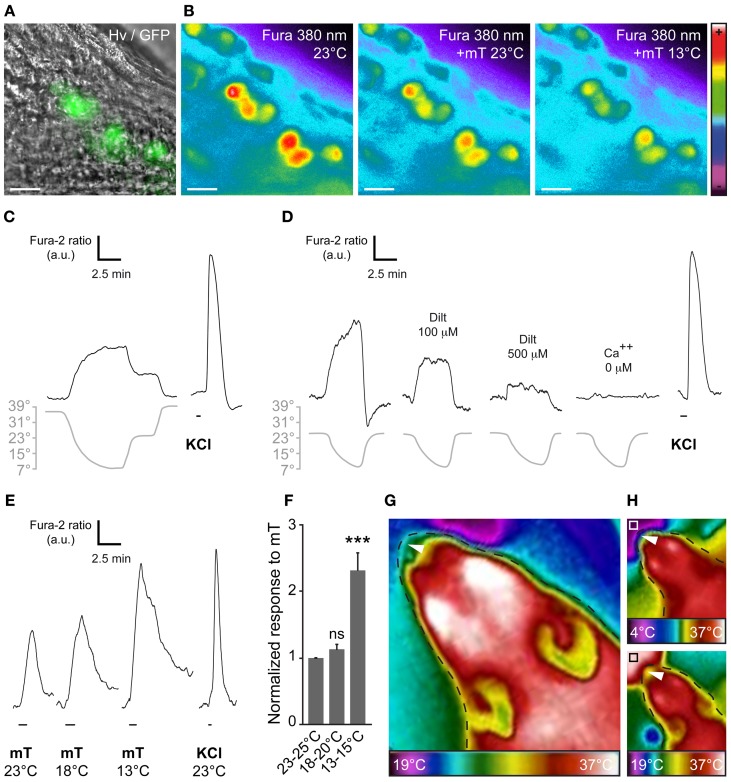
The mouse Grueneberg ganglion (GG) is an olfactory subsystem located at the tip of the nose close to the entry of the naris. It comprises neurons that are both sensitive to cold temperature and play an important role in the detection of alarm pheromones (APs). This chemical modality may be essential for species survival. Interestingly, GG neurons display an atypical mammalian olfactory morphology with neurons bearing deeply invaginated cilia mostly covered by ensheathing glial cells. We had previously noticed their morphological resemblance with the chemosensory amphid neurons found in the anterior region of the head of Caenorhabditis elegans (C. elegans). We demonstrate here further molecular and functional similarities. Thus, we found an orthologous expression of molecular signaling elements that was furthermore restricted to similar specific subcellular localizations. Calcium imaging also revealed a ligand selectivity for the methylated thiazole odorants that amphid neurons are known to detect. Cellular responses from GG neurons evoked by chemical or temperature stimuli were also partially cGMP-dependent. In addition, we found that, although behaviors depending on temperature sensing in the mouse, such as huddling and thermotaxis did not implicate the GG, the thermosensitivity modulated the chemosensitivity at the level of single GG neurons. Thus, the striking similarities with the chemosensory amphid neurons of C. elegans conferred to the mouse GG neurons unique multimodal sensory properties.
Keywords: Grueneberg ganglion; alarm pheromone; amphid neurons; behavior; calcium imaging; olfactory; temperature sensing.
Figures
Similar articles
-
Morphological and physiological species-dependent characteristics of the rodent Grueneberg ganglion.Front Neuroanat. 2014 Aug 27;8:87. doi: 10.3389/fnana.2014.00087. eCollection 2014. Front Neuroanat. 2014. PMID: 25221478 Free PMC article.
-
Expression of cGMP signaling elements in the Grueneberg ganglion.Histochem Cell Biol. 2009 Jan;131(1):75-88. doi: 10.1007/s00418-008-0514-8. Epub 2008 Oct 1. Histochem Cell Biol. 2009. PMID: 18830617
-
The Grueneberg ganglion: signal transduction and coding in an olfactory and thermosensory organ involved in the detection of alarm pheromones and predator-secreted kairomones.Cell Tissue Res. 2021 Jan;383(1):535-548. doi: 10.1007/s00441-020-03380-w. Epub 2021 Jan 6. Cell Tissue Res. 2021. PMID: 33404842 Review.
-
Grueneberg ganglion cells mediate alarm pheromone detection in mice.Science. 2008 Aug 22;321(5892):1092-5. doi: 10.1126/science.1160770. Science. 2008. PMID: 18719286
-
The Grueneberg ganglion: a novel sensory system in the nose.Histol Histopathol. 2010 Jul;25(7):909-15. doi: 10.14670/HH-25.909. Histol Histopathol. 2010. PMID: 20503179 Review.
Cited by
-
Olfactory memory networks: from emotional learning to social behaviors.Front Behav Neurosci. 2015 Feb 17;9:36. doi: 10.3389/fnbeh.2015.00036. eCollection 2015. Front Behav Neurosci. 2015. PMID: 25741259 Free PMC article. No abstract available.
-
The Grueneberg ganglion controls odor-driven food choices in mice under threat.Commun Biol. 2020 Sep 24;3(1):533. doi: 10.1038/s42003-020-01257-w. Commun Biol. 2020. PMID: 32973323 Free PMC article.
-
Alarm pheromone and kairomone detection via bitter taste receptors in the mouse Grueneberg ganglion.BMC Biol. 2018 Jan 18;16(1):12. doi: 10.1186/s12915-017-0479-y. BMC Biol. 2018. PMID: 29347925 Free PMC article.
-
Age-dependent appearance of SARS-CoV-2 entry sites in mouse chemosensory systems reflects COVID-19 anosmia-ageusia symptoms.Commun Biol. 2021 Jul 15;4(1):880. doi: 10.1038/s42003-021-02410-9. Commun Biol. 2021. PMID: 34267318 Free PMC article.
-
From In Vitro Data to In Vivo Interspecies Danger Communication: A Study of Chemosensing via the Mouse Grueneberg Ganglion.Animals (Basel). 2022 Feb 1;12(3):356. doi: 10.3390/ani12030356. Animals (Basel). 2022. PMID: 35158677 Free PMC article.
References
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous