

(13)C NMR spectroscopy applications to brain energy metabolism
- PMID: 24367329
- PMCID: PMC3856424
- DOI: 10.3389/fnene.2013.00009
(13)C NMR spectroscopy applications to brain energy metabolism
Abstract
(13)C nuclear magnetic resonance (NMR) spectroscopy is the method of choice for studying brain metabolism. Indeed, the most convincing data obtained to decipher metabolic exchanges between neurons and astrocytes have been obtained using this technique, thus illustrating its power. It may be difficult for non-specialists, however, to grasp thefull implication of data presented in articles written by spectroscopists. The aim of the review is, therefore, to provide a fundamental understanding of this topic to facilitate the non-specialists in their reading of this literature. In the first part of this review, we present the metabolic fate of (13)C-labeled substrates in the brain in a detailed way, including an overview of some general neurochemical principles. We also address and compare the various spectroscopic strategies that can be used to study brain metabolism. Then, we provide an overview of the (13)C NMR experiments performed to analyze both intracellular and intercellular metabolic fluxes. More particularly, the role of lactate as a potential energy substrate for neurons is discussed in the light of (13)C NMR data. Finally, new perspectives and applications offered by (13)C hyperpolarization are described.
Keywords: 13C NMR spectroscopy; astrocyte; brain metabolism; hyperpolarized NMR; metabolic modeling; neuroglial coupling; neuron.
Figures
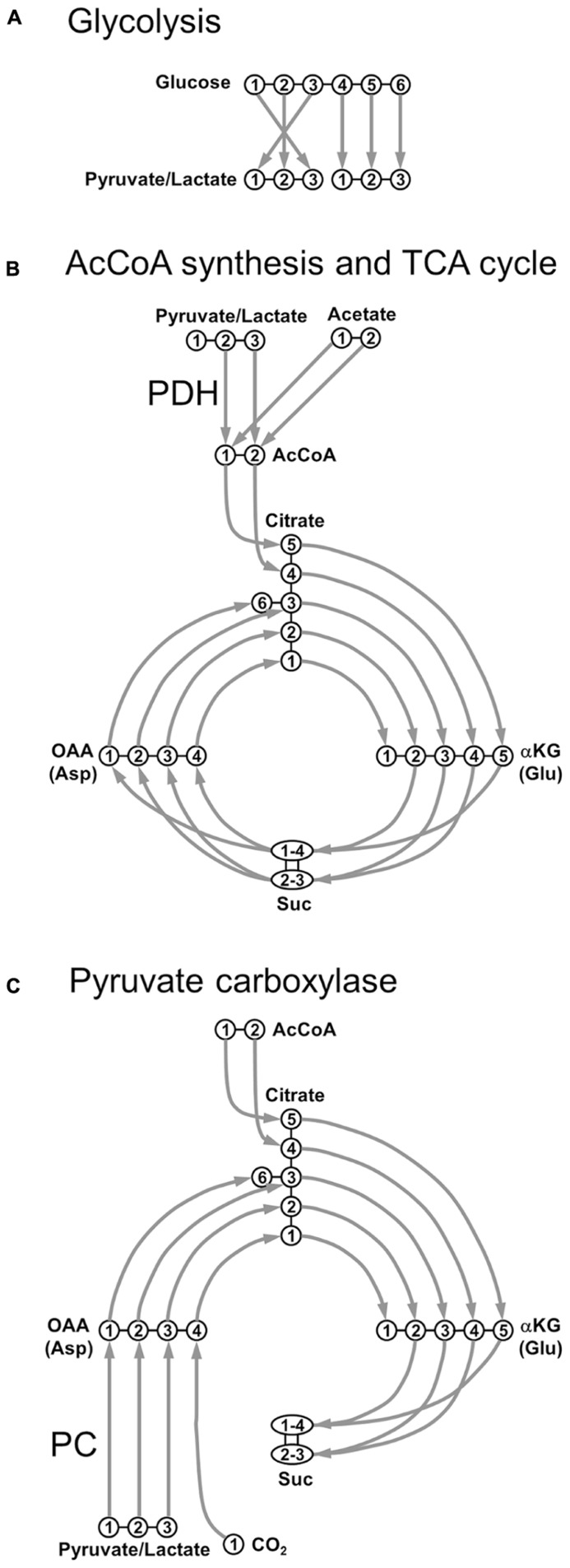
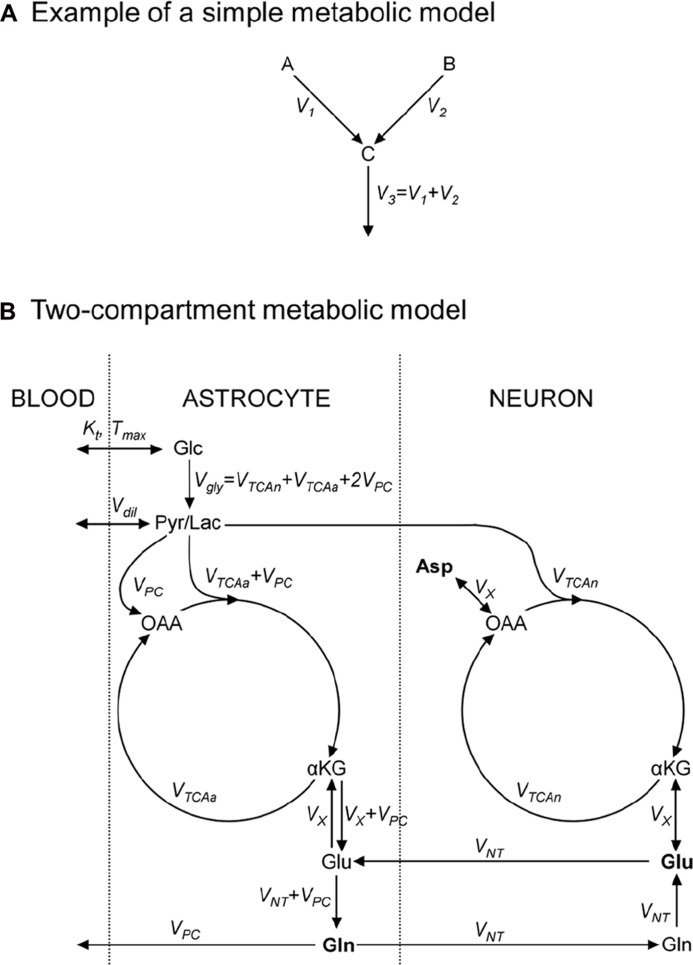
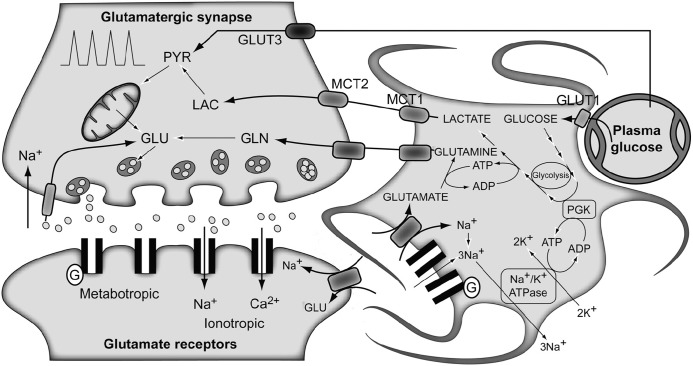
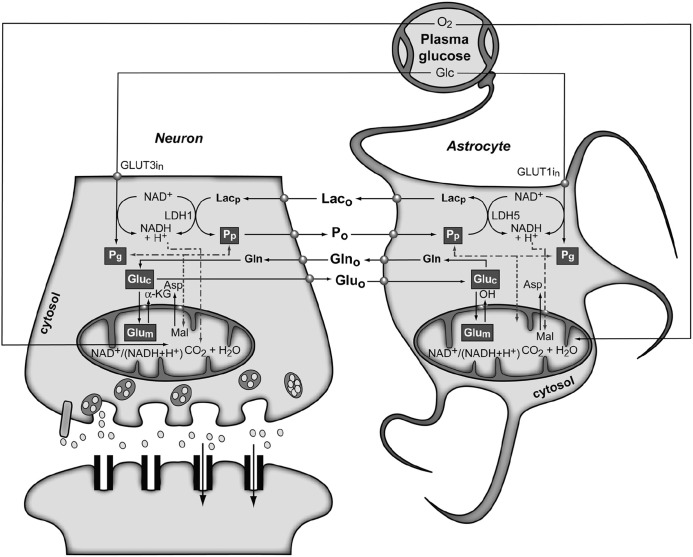
References
-
- Bachelard H. S. (1989). “Measurement of carbohydrates and their derivatives in neuronal tissues,” in Carbohydrates and Energy Metabolism eds Boulton A. A., Baker G. B., Butterworth R. F. (Clifton, NJ: Humana Press; ) 133–154
Publication types
LinkOut - more resources
Full Text Sources
Other Literature Sources