

The role of glutamine oxoglutarate aminotransferase and glutamate dehydrogenase in nitrogen metabolism in Mycobacterium bovis BCG
- PMID: 24367660
- PMCID: PMC3868603
- DOI: 10.1371/journal.pone.0084452
The role of glutamine oxoglutarate aminotransferase and glutamate dehydrogenase in nitrogen metabolism in Mycobacterium bovis BCG
Abstract
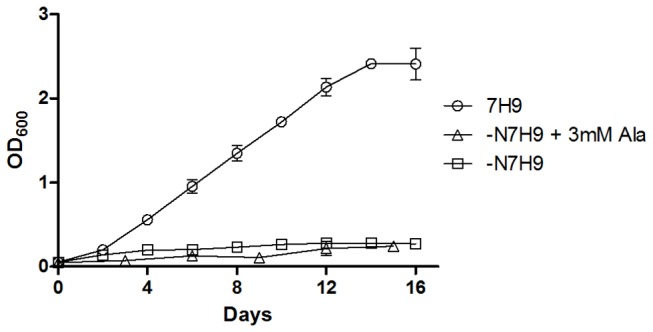
Recent evidence suggests that the regulation of intracellular glutamate levels could play an important role in the ability of pathogenic slow-growing mycobacteria to grow in vivo. However, little is known about the in vitro requirement for the enzymes which catalyse glutamate production and degradation in the slow-growing mycobacteria, namely; glutamine oxoglutarate aminotransferase (GOGAT) and glutamate dehydrogenase (GDH), respectively. We report that allelic replacement of the Mycobacterium bovis BCG gltBD-operon encoding for the large (gltB) and small (gltD) subunits of GOGAT with a hygromycin resistance cassette resulted in glutamate auxotrophy and that deletion of the GDH encoding-gene (gdh) led to a marked growth deficiency in the presence of L-glutamate as a sole nitrogen source as well as reduction in growth when cultured in an excess of L-asparagine.
Conflict of interest statement
Figures




Similar articles
-
Glutamate Dehydrogenase Is Required by Mycobacterium bovis BCG for Resistance to Cellular Stress.PLoS One. 2016 Jan 29;11(1):e0147706. doi: 10.1371/journal.pone.0147706. eCollection 2016. PLoS One. 2016. PMID: 26824899 Free PMC article.
-
Inorganic nitrogen assimilation in yeasts: alteration in enzyme activities associated with changes in cultural conditions and growth phase.J Bacteriol. 1972 Jan;109(1):25-33. doi: 10.1128/jb.109.1.25-33.1972. J Bacteriol. 1972. PMID: 4400414 Free PMC article.
-
Glutamine synthetase-glutamate synthase pathway and glutamate dehydrogenase play distinct roles in the sink-source nitrogen cycle in tobacco.Plant Physiol. 2006 Feb;140(2):444-56. doi: 10.1104/pp.105.071910. Epub 2006 Jan 11. Plant Physiol. 2006. PMID: 16407450 Free PMC article.
-
An Overview of Important Enzymes Involved in Nitrogen Assimilation of Plants.Methods Mol Biol. 2020;2057:1-13. doi: 10.1007/978-1-4939-9790-9_1. Methods Mol Biol. 2020. PMID: 31595465 Review.
-
Glutamate oxidation in astrocytes: Roles of glutamate dehydrogenase and aminotransferases.J Neurosci Res. 2016 Dec;94(12):1561-1571. doi: 10.1002/jnr.23908. Epub 2016 Sep 15. J Neurosci Res. 2016. PMID: 27629247 Review.
Cited by
-
Mycobacterium tuberculosis in the Face of Host-Imposed Nutrient Limitation.Microbiol Spectr. 2017 Jun;5(3):10.1128/microbiolspec.tbtb2-0030-2016. doi: 10.1128/microbiolspec.TBTB2-0030-2016. Microbiol Spectr. 2017. PMID: 28597811 Free PMC article. Review.
-
Nitrogen metabolism in Mycobacterium tuberculosis physiology and virulence.Nat Rev Microbiol. 2014 Nov;12(11):729-37. doi: 10.1038/nrmicro3349. Epub 2014 Sep 22. Nat Rev Microbiol. 2014. PMID: 25244084 Review.
-
Prominent transcriptomic changes in Mycobacterium intracellulare under acidic and oxidative stress.BMC Genomics. 2024 Apr 17;25(1):376. doi: 10.1186/s12864-024-10292-4. BMC Genomics. 2024. PMID: 38632539 Free PMC article.
-
Glutamate Dehydrogenase Is Required by Mycobacterium bovis BCG for Resistance to Cellular Stress.PLoS One. 2016 Jan 29;11(1):e0147706. doi: 10.1371/journal.pone.0147706. eCollection 2016. PLoS One. 2016. PMID: 26824899 Free PMC article.
-
PknG senses amino acid availability to control metabolism and virulence of Mycobacterium tuberculosis.PLoS Pathog. 2017 May 17;13(5):e1006399. doi: 10.1371/journal.ppat.1006399. eCollection 2017 May. PLoS Pathog. 2017. PMID: 28545104 Free PMC article.
References
-
- Harth G, Horwitz MA (1997) Expression and efficient export of enzymatically active Mycobacterium tuberculosis glutamine synthetase in Mycobacterium smegmatis and evidence that the information for export is contained within the protein. J Biol Chem 272: 22728–22735. doi:10.1074/jbc.272.36.22728. PubMed: 9278431. - DOI - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
