

Iron in intracellular infection: to provide or to deprive?
- PMID: 24367768
- PMCID: PMC3856365
- DOI: 10.3389/fcimb.2013.00096
Iron in intracellular infection: to provide or to deprive?
Abstract
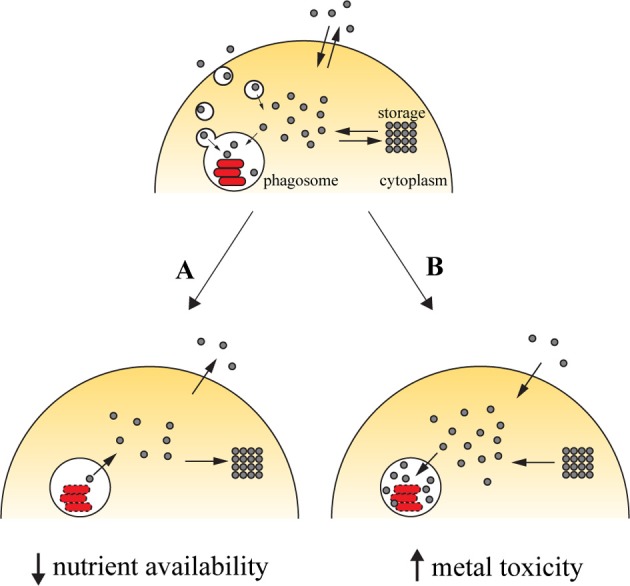
Due to their chemical versatility, transition metals were incorporated as cofactors for several basic metabolic pathways in living organisms. This same characteristic makes them potentially harmful, since they can be engaged in deleterious reactions like Fenton chemistry. As such, organisms have evolved highly specialized mechanisms to supply their own metal needs while keeping their toxic potential in check. This dual character comes into play in host-pathogen interactions, given that the host can either deprive the pathogen of these key nutrients or exploit them to induce toxicity toward the invading agent. Iron stands as the prototypic example of how a metal can be used to limit the growth of pathogens by nutrient deprivation, a mechanism widely studied in Mycobacterium infections. However, the host can also take advantage of iron-induced toxicity to control pathogen proliferation, as observed in infections caused by Leishmania. Whether we may harness either of the two pathways for therapeutical purposes is still ill-defined. In this review, we discuss how modulation of the host iron availability impacts the course of infections, focusing on those caused by two relevant intracellular pathogens, Mycobacterium and Leishmania.
Keywords: Leishmania; Mycobacterium; immunity; infection; transition metal.
Figures
Similar articles
-
Iron acquisition in Leishmania and its crucial role in infection.Parasitology. 2016 Sep;143(11):1347-57. doi: 10.1017/S0031182016000858. Epub 2016 May 25. Parasitology. 2016. PMID: 27221985 Review.
-
Leishmania infection triggers hepcidin-mediated proteasomal degradation of Nramp1 to increase phagolysosomal iron availability.Cell Microbiol. 2020 Dec;22(12):e13253. doi: 10.1111/cmi.13253. Epub 2020 Sep 10. Cell Microbiol. 2020. PMID: 32827218
-
Role of divalent metals in infectious disease susceptibility and outcome.Clin Microbiol Infect. 2018 Jan;24(1):16-23. doi: 10.1016/j.cmi.2017.01.018. Epub 2017 Jan 29. Clin Microbiol Infect. 2018. PMID: 28143784 Review.
-
Metal limitation and toxicity at the interface between host and pathogen.FEMS Microbiol Rev. 2014 Nov;38(6):1235-49. doi: 10.1111/1574-6976.12087. Epub 2014 Sep 29. FEMS Microbiol Rev. 2014. PMID: 25211180 Free PMC article. Review.
-
The struggle for iron - a metal at the host-pathogen interface.Cell Microbiol. 2010 Dec;12(12):1691-702. doi: 10.1111/j.1462-5822.2010.01529.x. Epub 2010 Oct 21. Cell Microbiol. 2010. PMID: 20964797 Review.
Cited by
-
Manipulation of the mononuclear phagocyte system by Mycobacterium tuberculosis.Cold Spring Harb Perspect Med. 2014 Aug 21;4(11):a018549. doi: 10.1101/cshperspect.a018549. Cold Spring Harb Perspect Med. 2014. PMID: 25147188 Free PMC article. Review.
-
Survival in Hostile Conditions: Pupylation and the Proteasome in Actinobacterial Stress Response Pathways.Front Mol Biosci. 2021 Jun 7;8:685757. doi: 10.3389/fmolb.2021.685757. eCollection 2021. Front Mol Biosci. 2021. PMID: 34179091 Free PMC article. Review.
-
Metal economy in host-microbe interactions.Front Cell Infect Microbiol. 2015 Jan 13;4:190. doi: 10.3389/fcimb.2014.00190. eCollection 2014. Front Cell Infect Microbiol. 2015. PMID: 25629009 Free PMC article. No abstract available.
-
TH1/TH2 paradigm extended: macrophage polarization as an unappreciated pathogen-driven escape mechanism?Front Immunol. 2014 Nov 26;5:603. doi: 10.3389/fimmu.2014.00603. eCollection 2014. Front Immunol. 2014. PMID: 25505468 Free PMC article. Review.
-
Advances in Ferritin Physiology and Possible Implications in Bacterial Infection.Int J Mol Sci. 2023 Feb 28;24(5):4659. doi: 10.3390/ijms24054659. Int J Mol Sci. 2023. PMID: 36902088 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials