

H3K4 mono- and di-methyltransferase MLL4 is required for enhancer activation during cell differentiation
- PMID: 24368734
- PMCID: PMC3869375
- DOI: 10.7554/eLife.01503
H3K4 mono- and di-methyltransferase MLL4 is required for enhancer activation during cell differentiation
Abstract
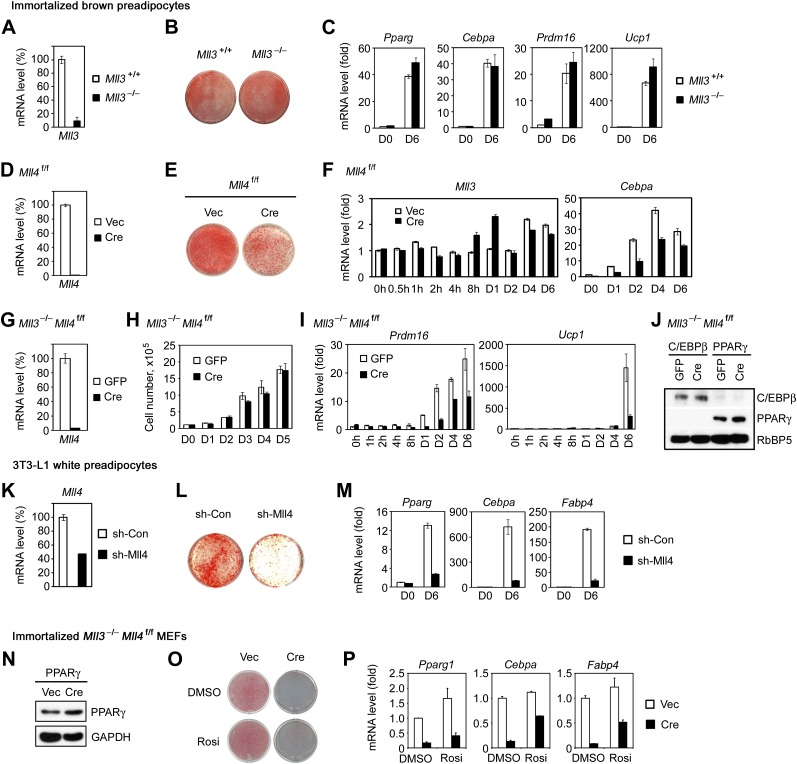
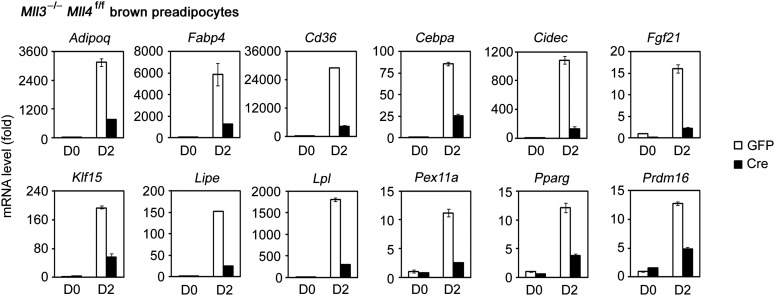
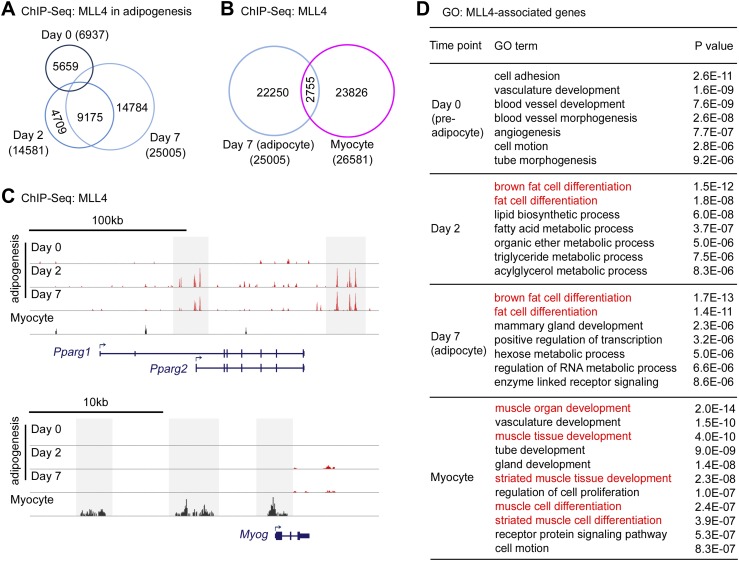
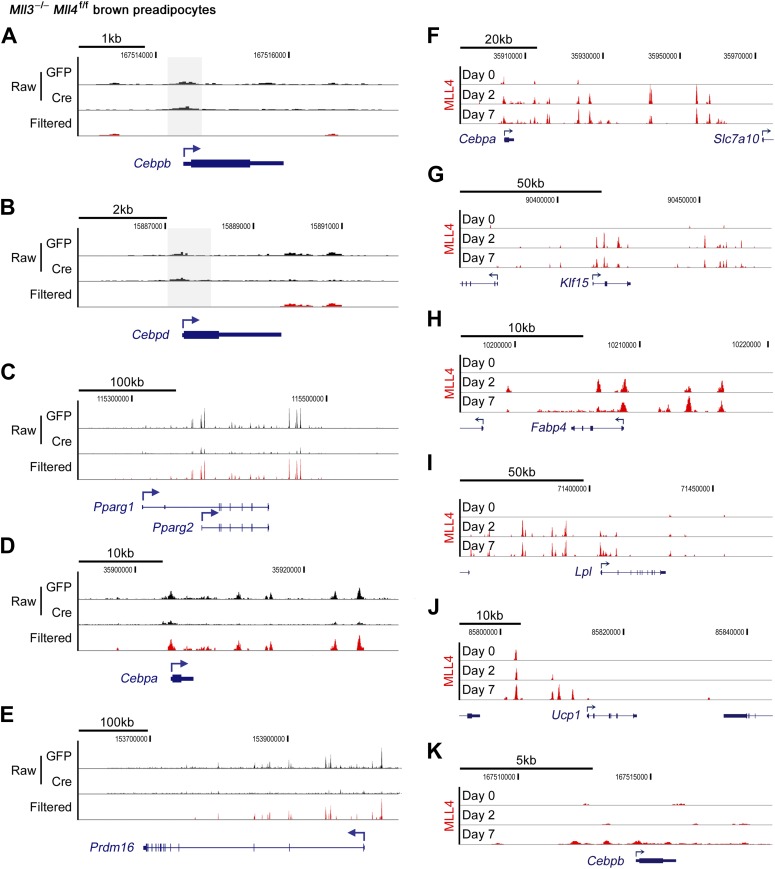
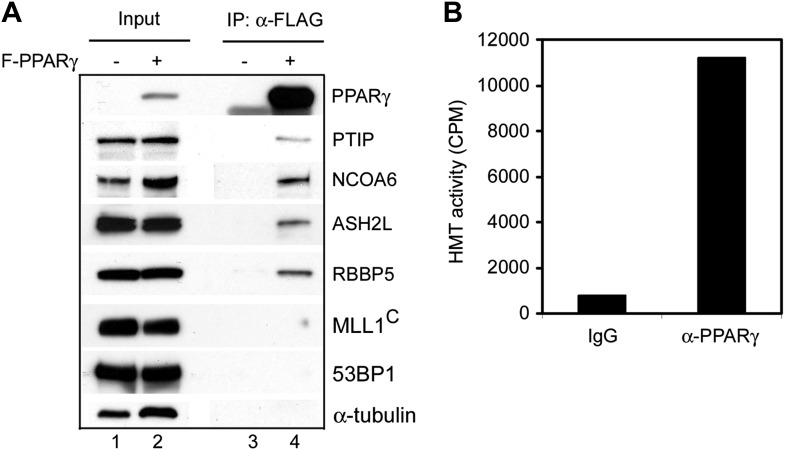
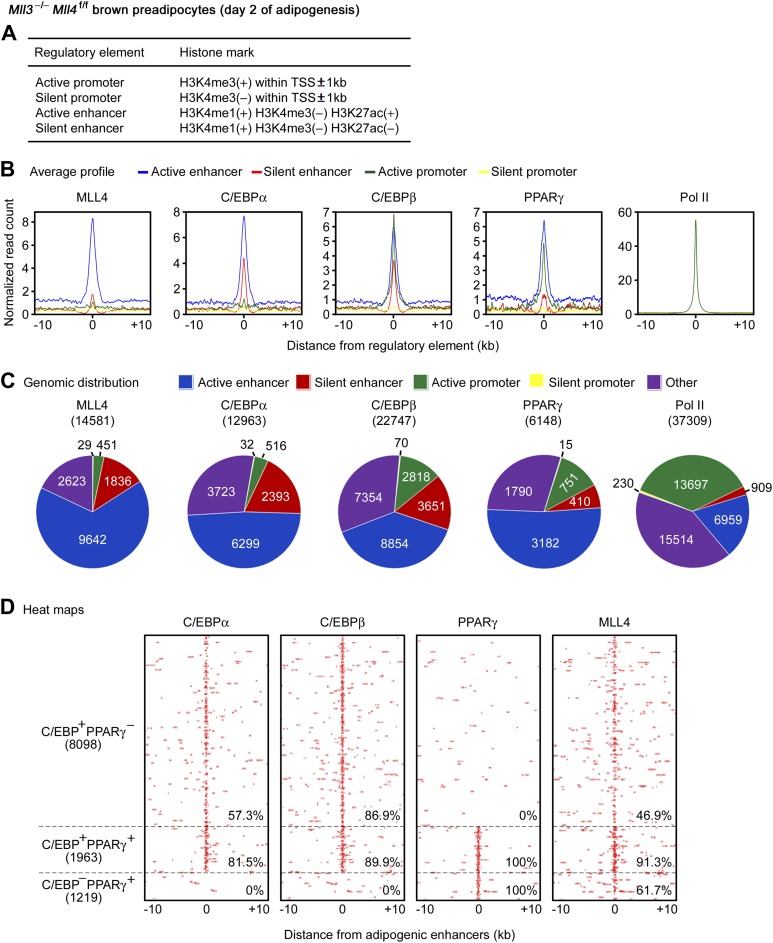
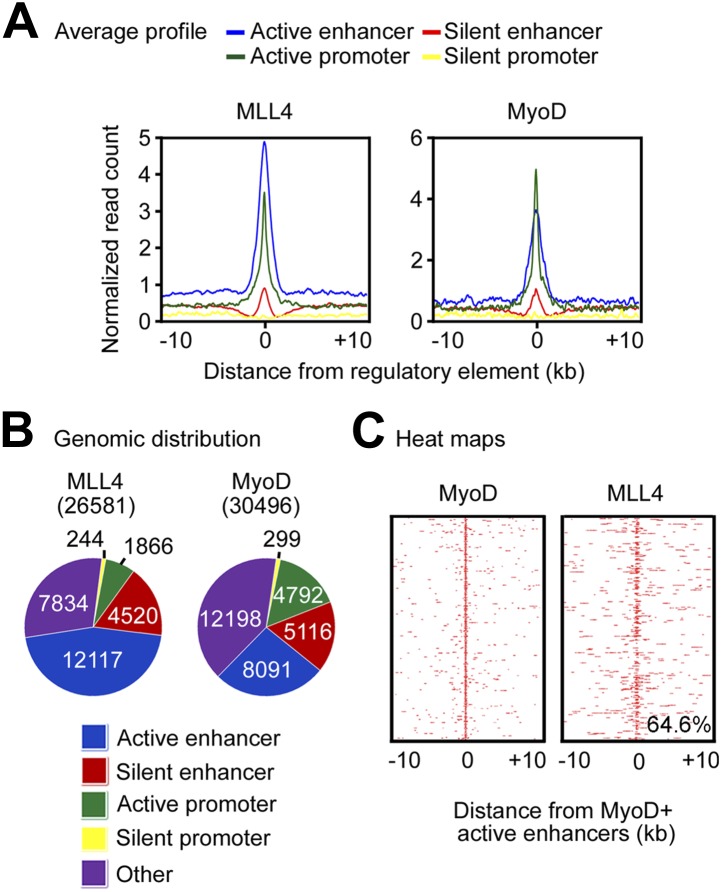
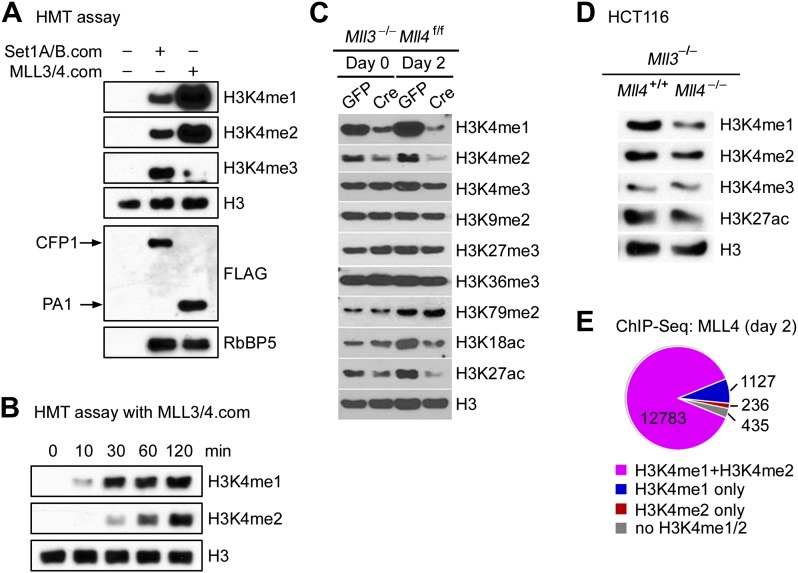
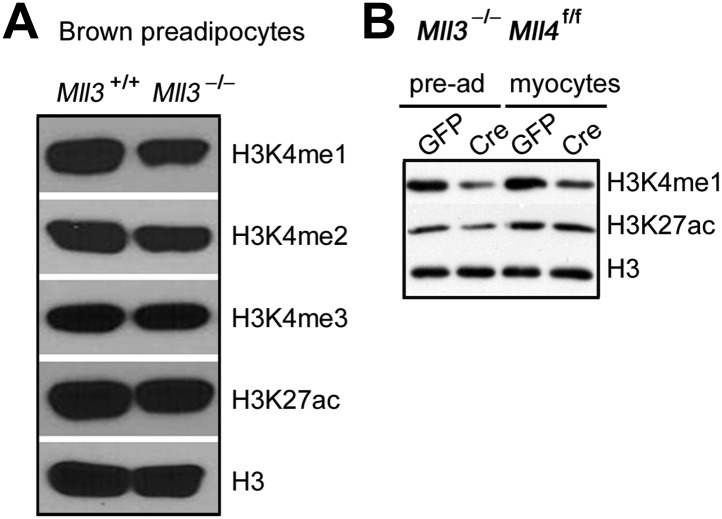
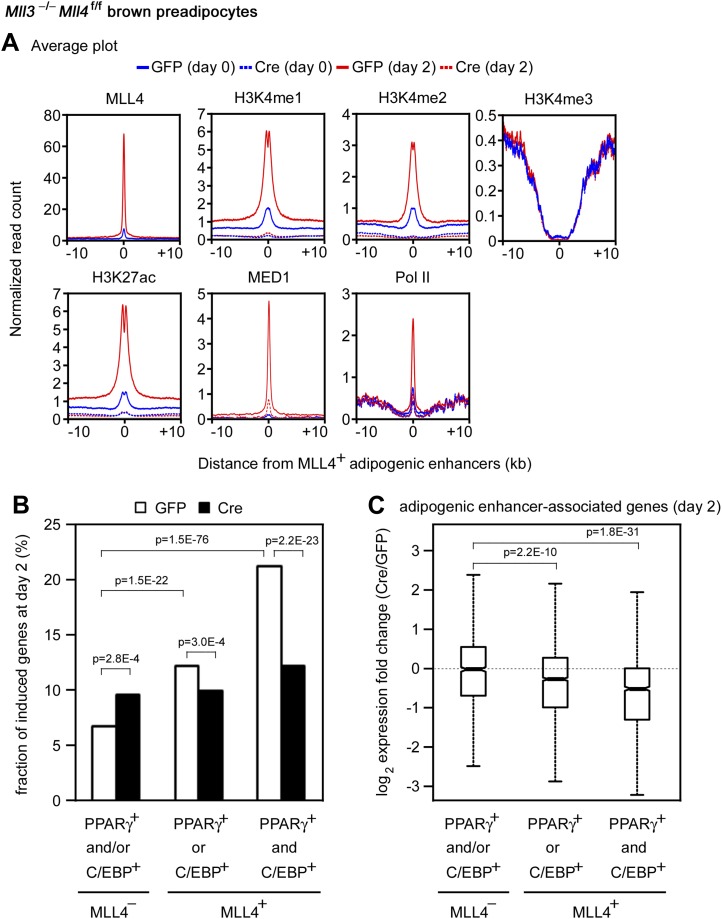
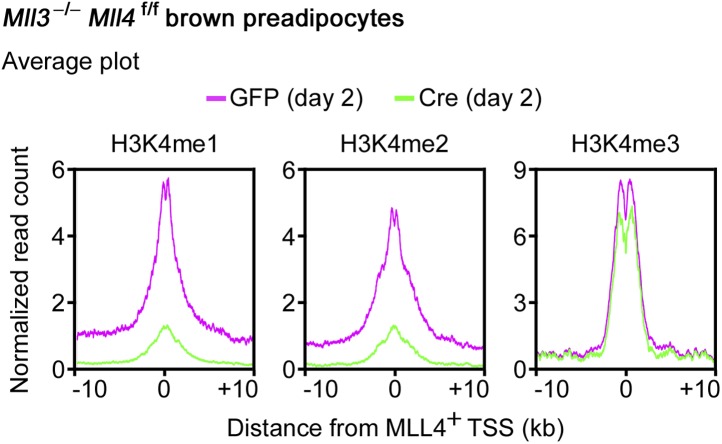
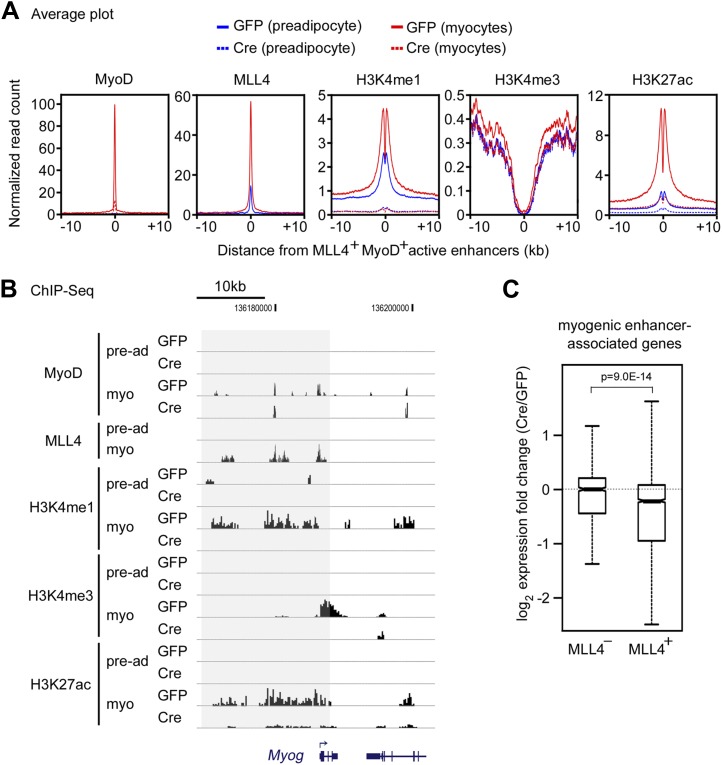
Enhancers play a central role in cell-type-specific gene expression and are marked by H3K4me1/2. Active enhancers are further marked by H3K27ac. However, the methyltransferases responsible for H3K4me1/2 on enhancers remain elusive. Furthermore, how these enzymes function on enhancers to regulate cell-type-specific gene expression is unclear. In this study, we identify MLL4 (KMT2D) as a major mammalian H3K4 mono- and di-methyltransferase with partial functional redundancy with MLL3 (KMT2C). Using adipogenesis and myogenesis as model systems, we show that MLL4 exhibits cell-type- and differentiation-stage-specific genomic binding and is predominantly localized on enhancers. MLL4 co-localizes with lineage-determining transcription factors (TFs) on active enhancers during differentiation. Deletion of Mll4 markedly decreases H3K4me1/2, H3K27ac, Mediator and Polymerase II levels on enhancers and leads to severe defects in cell-type-specific gene expression and cell differentiation. Together, these findings identify MLL4 as a major mammalian H3K4 mono- and di-methyltransferase essential for enhancer activation during cell differentiation. DOI: http://dx.doi.org/10.7554/eLife.01503.001.
Keywords: H3K27ac; H3K4me1; KMT2D; MLL4; adipogenesis; cell differentiation; enhancer activation; enhancer chromatin modification; myogenesis.
Conflict of interest statement
The authors declare that no competing interests exist.
Figures
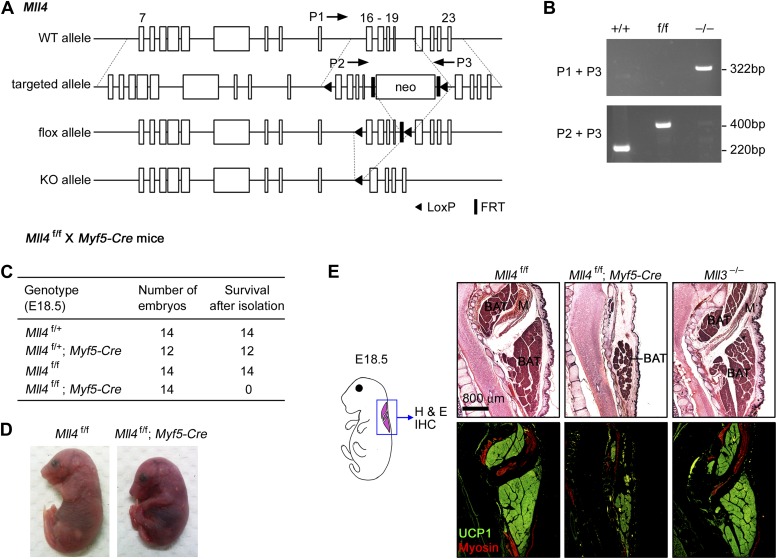
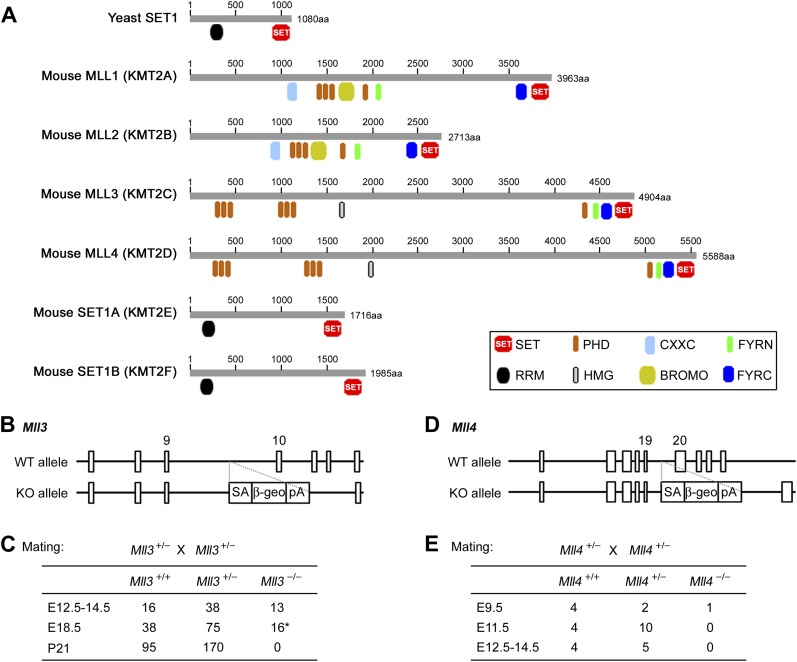
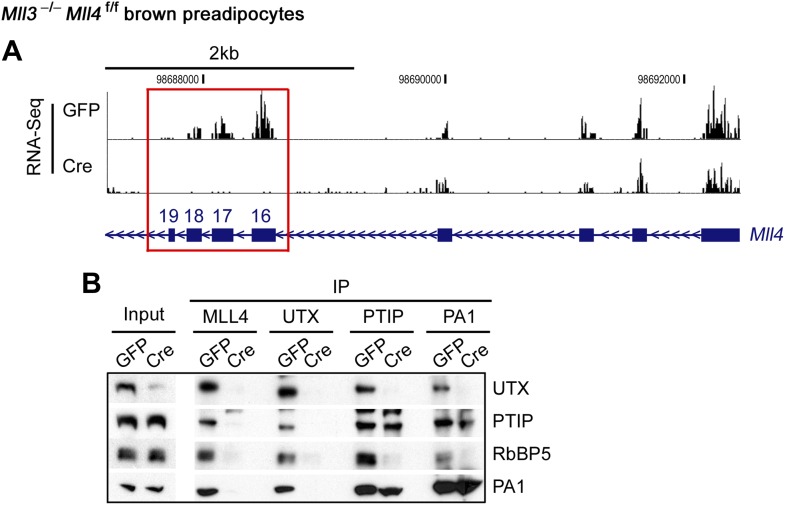
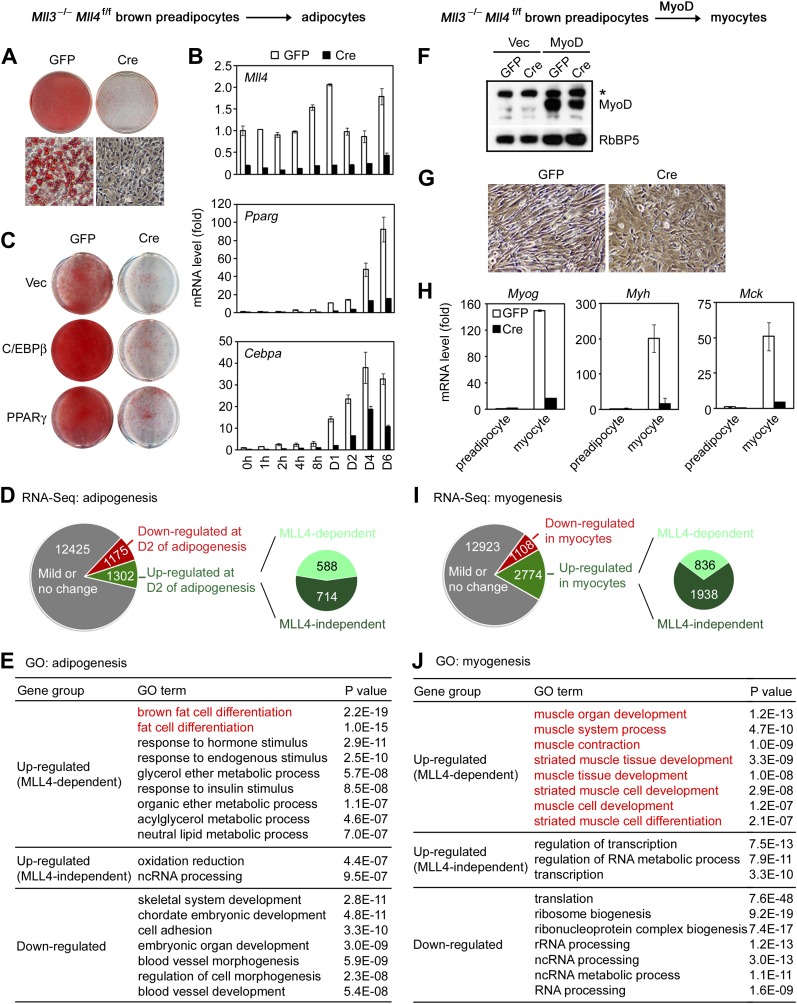
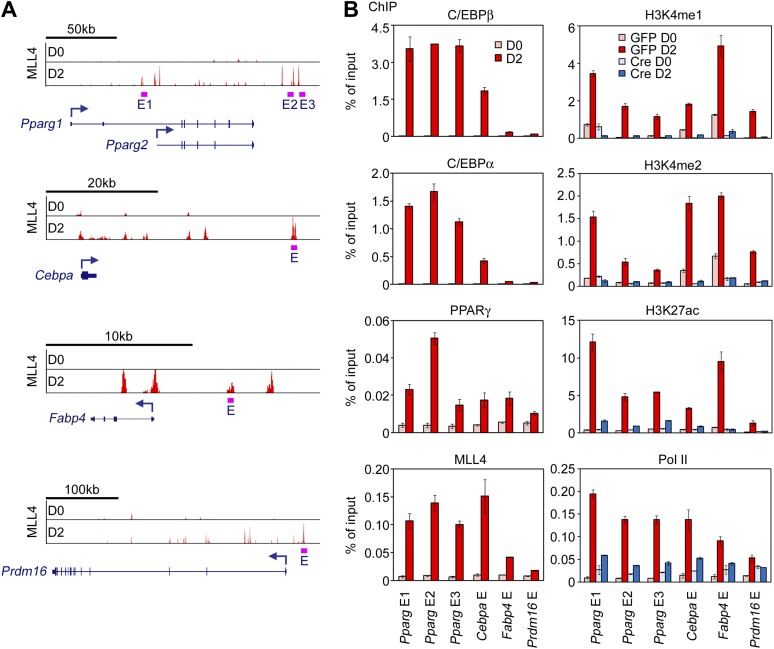
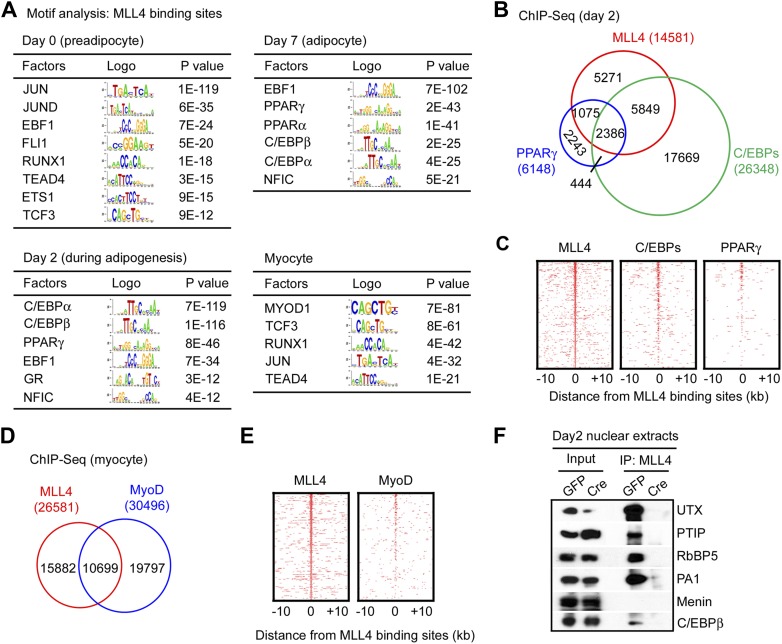
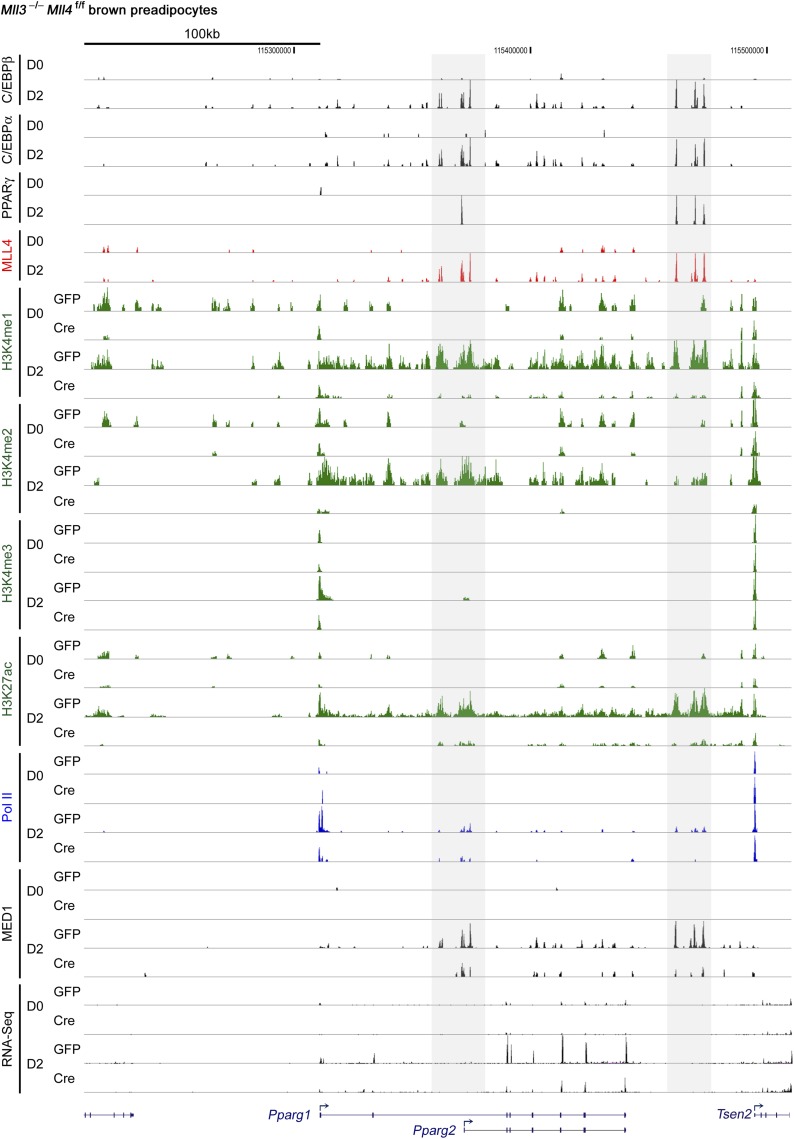
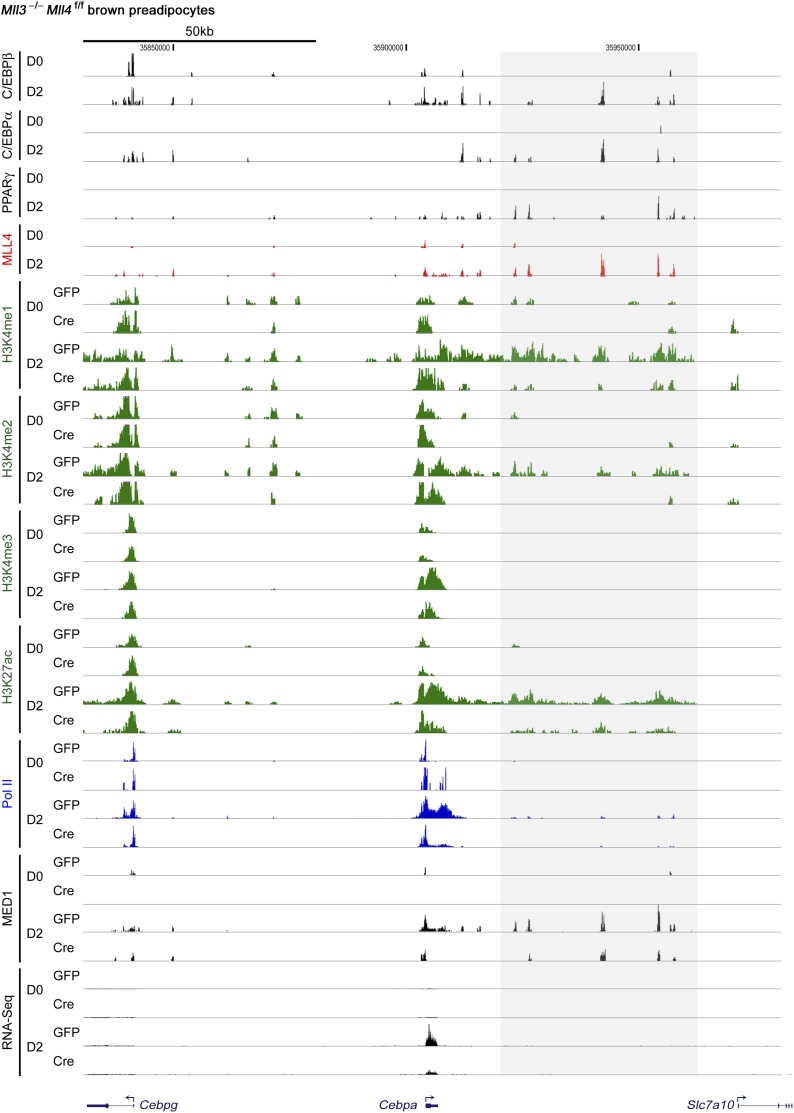
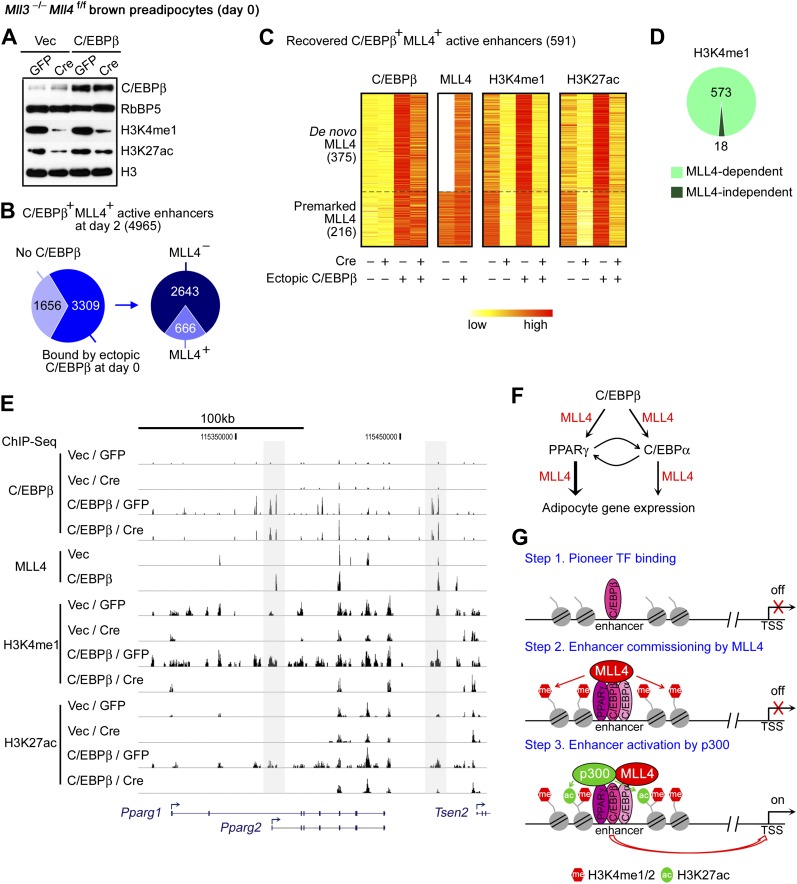
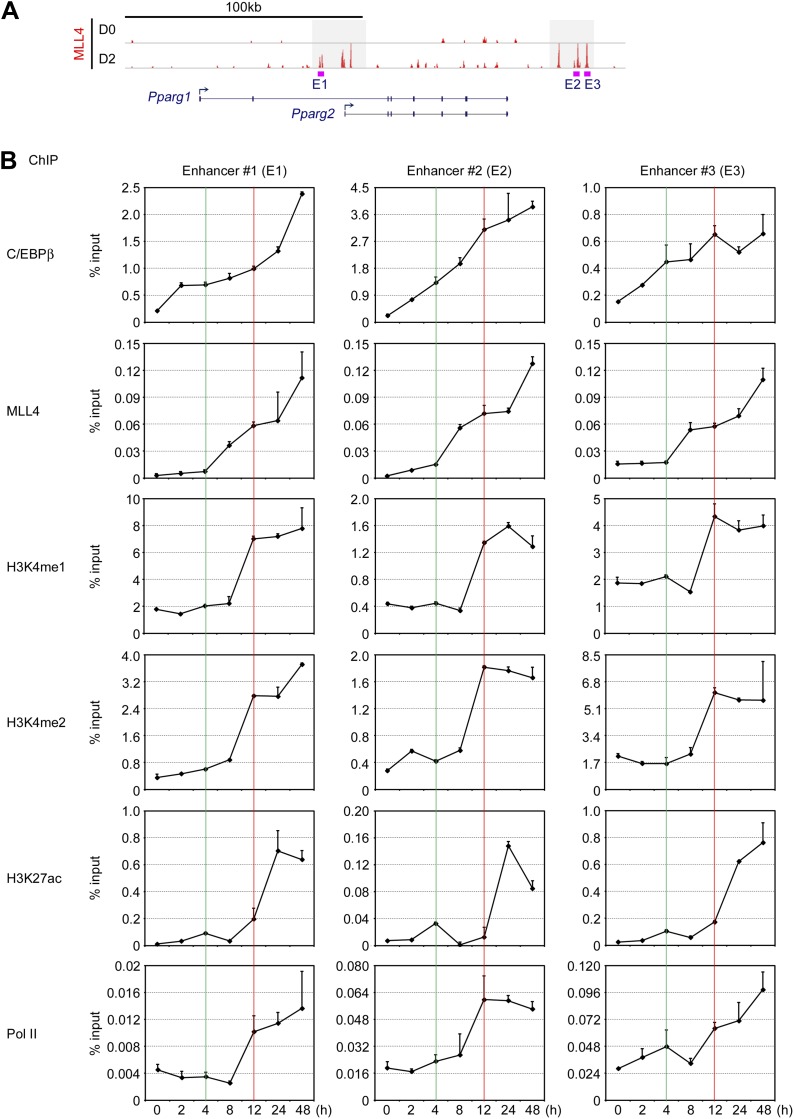
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
