

Interferon-inducible protein IFI35 negatively regulates RIG-I antiviral signaling and supports vesicular stomatitis virus replication
- PMID: 24371060
- PMCID: PMC3957919
- DOI: 10.1128/JVI.03202-13
Interferon-inducible protein IFI35 negatively regulates RIG-I antiviral signaling and supports vesicular stomatitis virus replication
Abstract
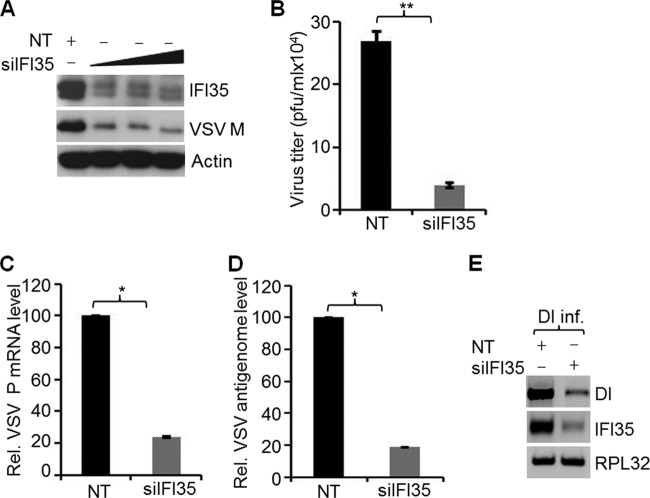
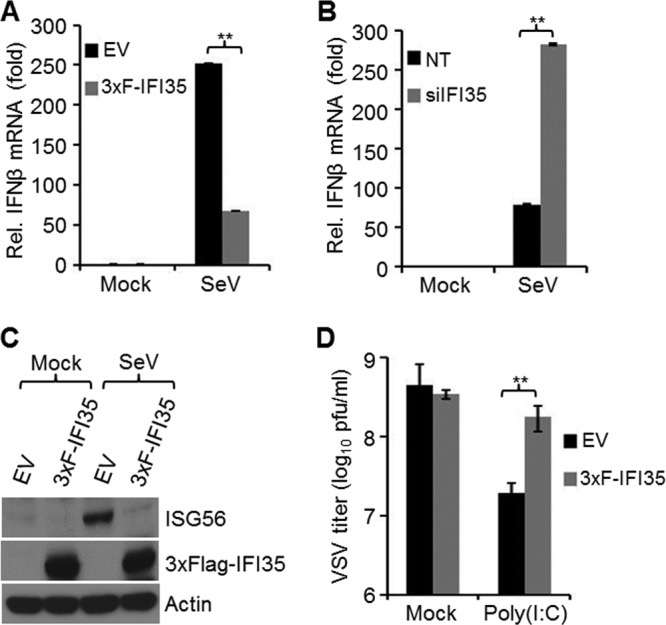
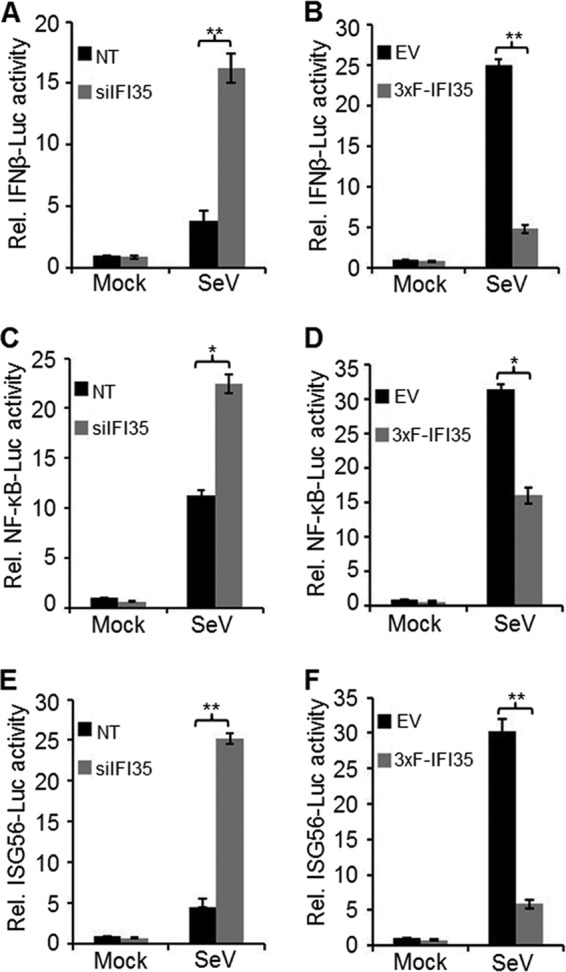
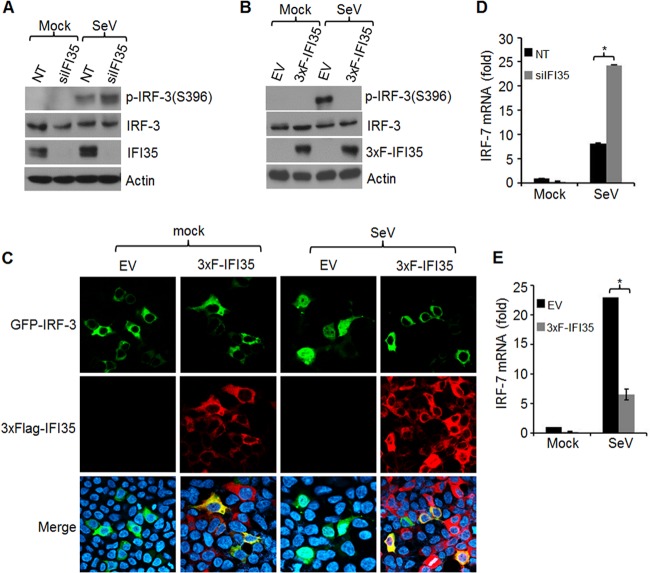
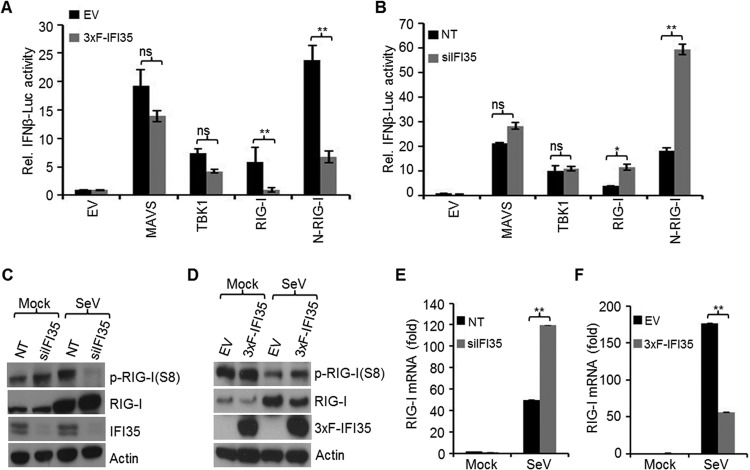
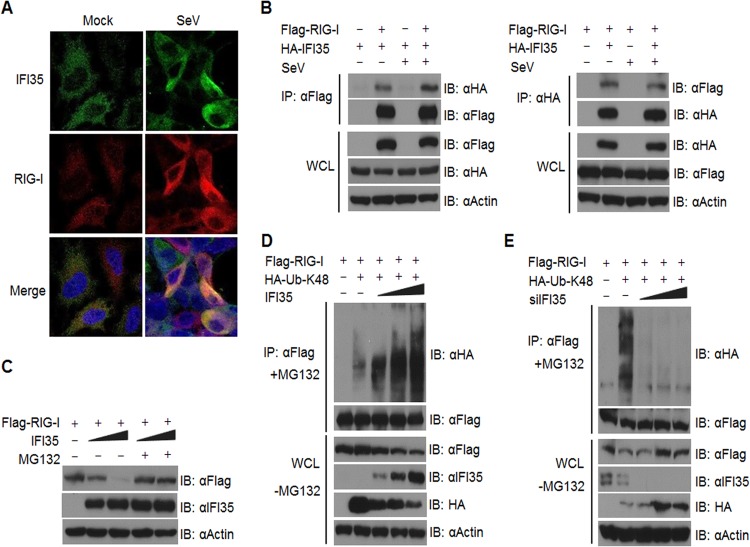
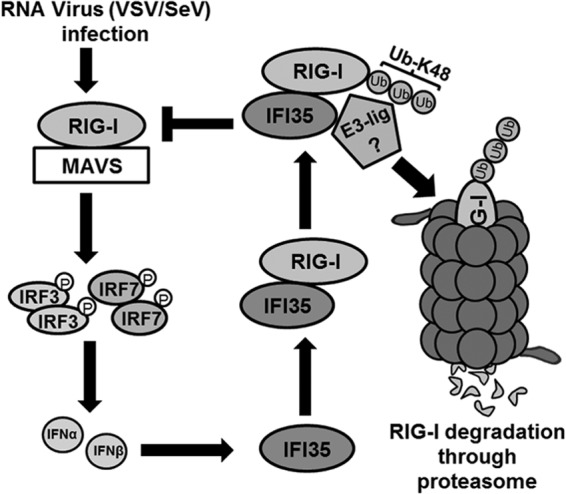
In a genome-wide small interfering RNA (siRNA) screen, we recently identified the interferon (IFN)-inducible protein 35 (IFI35; also known as IFP35) as a factor required for vesicular stomatitis virus (VSV) infection. Studies reported here were conducted to further understand the role and requirement of IFI35 in VSV infection. Consistent with the siRNA screening data, we found that depletion of IFI35 led to reduced VSV replication at the level of viral gene expression. Although no direct interaction of IFI35 with the viral replication machinery was observed, we found that IFI35 negatively regulated the host innate immune response and rescued poly(I·C)-induced inhibition of VSV replication. Promoter-driven reporter gene assays demonstrated that IFI35 overexpression suppressed the activation of IFN-β and ISG56 promoters, whereas its depletion had the opposite effect. Further investigation revealed that IFI35 specifically interacted with retinoic acid-inducible gene I (RIG-I) and negatively regulated its activation through mechanisms that included (i) suppression of dephosphorylation (activation) of RIG-I and (ii) proteasome-mediated degradation of RIG-I via K48-linked ubiquitination. Overall, the results presented here suggest a novel role for IFI35 in negative regulation of RIG-I-mediated antiviral signaling, which will have implications for diseases associated with excessive immune signaling.
Importance: Mammalian cells employ a variety of mechanisms, including production of interferons (IFNs), to counteract invading pathogens. In this study, we identified a novel role for a cellular protein, IFN-inducible protein 35 (IFP35/IFI35), in negatively regulating the host IFN response during vesicular stomatitis virus (VSV) infection. Specifically, we found that IFI35 inhibited activation of the RNA sensor, the retinoic acid-inducible gene I (RIG-I), leading to inhibition of IFN production and thus resulting in better replication of VSV. The identification of a cellular factor that attenuates the IFN response will have implications toward understanding inflammatory diseases in humans that have been found to be associated with defects in the regulation of host IFN production, such as systemic lupus erythematosus and psoriasis.
Figures
References
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
