

Mutation of the palmitoylation site of estrogen receptor α in vivo reveals tissue-specific roles for membrane versus nuclear actions
- PMID: 24371309
- PMCID: PMC3896153
- DOI: 10.1073/pnas.1322057111
Mutation of the palmitoylation site of estrogen receptor α in vivo reveals tissue-specific roles for membrane versus nuclear actions
Abstract
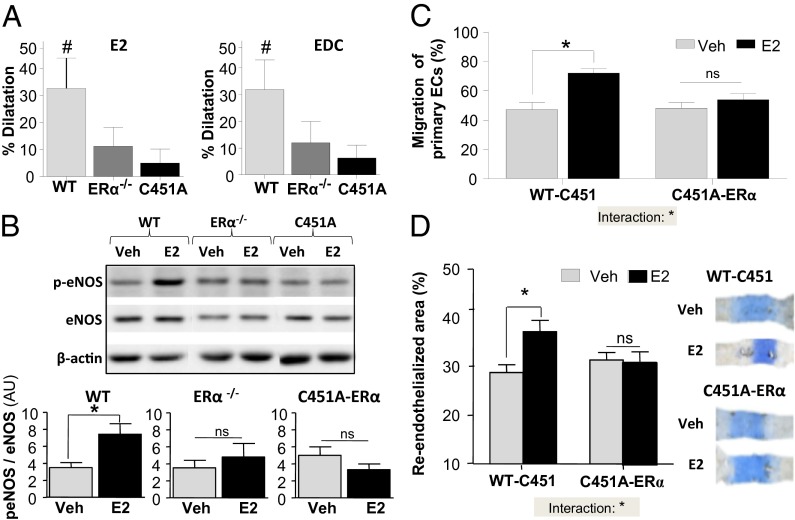
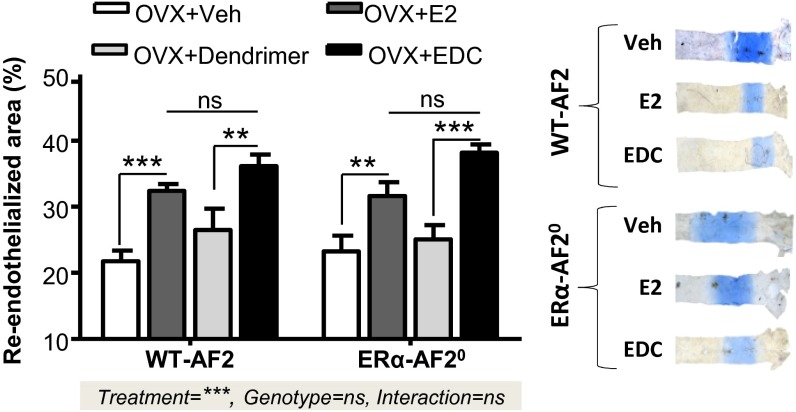
Estrogen receptor alpha (ERα) activation functions AF-1 and AF-2 classically mediate gene transcription in response to estradiol (E2). A fraction of ERα is targeted to plasma membrane and elicits membrane-initiated steroid signaling (MISS), but the physiological roles of MISS in vivo are poorly understood. We therefore generated a mouse with a point mutation of the palmitoylation site of ERα (C451A-ERα) to obtain membrane-specific loss of function of ERα. The abrogation of membrane localization of ERα in vivo was confirmed in primary hepatocytes, and it resulted in female infertility with abnormal ovaries lacking corpora lutea and increase in luteinizing hormone levels. In contrast, E2 action in the uterus was preserved in C451A-ERα mice and endometrial epithelial proliferation was similar to wild type. However, E2 vascular actions such as rapid dilatation, acceleration of endothelial repair, and endothelial NO synthase phosphorylation were abrogated in C451A-ERα mice. A complementary mutant mouse lacking the transactivation function AF-2 of ERα (ERα-AF2(0)) provided selective loss of function of nuclear ERα actions. In ERα-AF2(0), the acceleration of endothelial repair in response to estrogen-dendrimer conjugate, which is a membrane-selective ER ligand, was unaltered, demonstrating integrity of MISS actions. In genome-wide analysis of uterine gene expression, the vast majority of E2-dependent gene regulation was abrogated in ERα-AF2(0), whereas in C451A-ERα it was nearly fully preserved, indicating that membrane-to-nuclear receptor cross-talk in vivo is modest in the uterus. Thus, this work genetically segregated membrane versus nuclear actions of a steroid hormone receptor and demonstrated their in vivo tissue-specific roles.
Keywords: fertility; genomic actions; nongenomic effects; vascular effects.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Hewitt SC, Harrell JC, Korach KS. Lessons in estrogen biology from knockout and transgenic animals. Annu Rev Physiol. 2005;67:285–308. - PubMed
-
- Nilsson S, Koehler KF, Gustafsson JA. Development of subtype-selective oestrogen receptor-based therapeutics. Nat Rev Drug Discov. 2011;10(10):778–792. - PubMed
-
- Abot A, et al. The AF-1 activation function of estrogen receptor α is necessary and sufficient for uterine epithelial cell proliferation in vivo. Endocrinology. 2013;154(6):2222–2233. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
