

Quantitative genetics in the study of virus-induced disease
- PMID: 24373313
- PMCID: PMC7271172
- DOI: 10.1016/B978-0-12-800098-4.00004-0
Quantitative genetics in the study of virus-induced disease
Abstract
While the role of viral variants has long been known to play a key role in causing variation in disease severity, it is also clear that host genetic variation plays a critical role in determining virus-induced disease responses. However, a variety of factors, including confounding environmental variables, rare genetic variants requiring extremely large cohorts, the temporal dynamics of infections, and ethical limitation on human studies, have made the identification and dissection of variant host genes and pathways difficult within human populations. This difficulty has led to the development of a variety of experimental approaches used to identify host genetic contributions to disease responses. In this chapter, we describe the history of genetic associations within the human population, the development of experimentally tractable systems, and the insights these specific approaches provide. We conclude with a discussion of recent advances that allow for the investigation of the role of complex genetic networks that underlie host responses to infection, with the goal of drawing connections to human infections. In particular, we highlight the need for robust animal models with which to directly control and assess the role of host genetics on viral infection outcomes.
Keywords: Association; Complex trait; GWAS; Genetic mapping; Genetic reference population; Linkage; QTL; Viral disease.
© 2014 Elsevier Inc. All rights reserved.
Figures
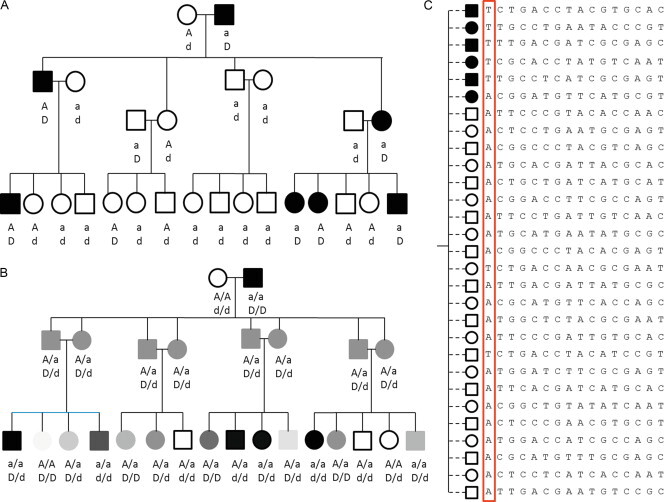
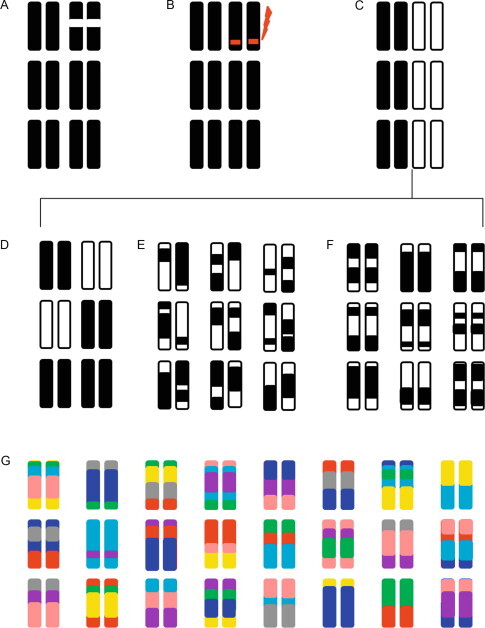
Similar articles
-
Dissection of Host Susceptibility to Bacterial Infections and Its Toxins.Methods Mol Biol. 2017;1488:551-578. doi: 10.1007/978-1-4939-6427-7_27. Methods Mol Biol. 2017. PMID: 27933544
-
Giving the Genes a Shuffle: Using Natural Variation to Understand Host Genetic Contributions to Viral Infections.Trends Genet. 2018 Oct;34(10):777-789. doi: 10.1016/j.tig.2018.07.005. Epub 2018 Aug 18. Trends Genet. 2018. PMID: 30131185 Free PMC article. Review.
-
Human Genetic Determinants of Viral Diseases.Annu Rev Genet. 2017 Nov 27;51:241-263. doi: 10.1146/annurev-genet-120116-023425. Epub 2017 Aug 30. Annu Rev Genet. 2017. PMID: 28853921 Free PMC article. Review.
-
Discovery of Variants Underlying Host Susceptibility to Virus Infection Using Whole-Exome Sequencing.Methods Mol Biol. 2017;1656:209-227. doi: 10.1007/978-1-4939-7237-1_14. Methods Mol Biol. 2017. PMID: 28808973 Free PMC article.
-
Systems Genetics as a Tool to Identify Master Genetic Regulators in Complex Disease.Methods Mol Biol. 2017;1488:337-362. doi: 10.1007/978-1-4939-6427-7_16. Methods Mol Biol. 2017. PMID: 27933533
Cited by
-
Chronic chikungunya virus musculoskeletal disease: what are the underlying mechanisms?Future Microbiol. 2016;11(3):331-4. doi: 10.2217/fmb.15.147. Epub 2016 Mar 4. Future Microbiol. 2016. PMID: 26939523 Free PMC article. No abstract available.
-
Oas1b-dependent Immune Transcriptional Profiles of West Nile Virus Infection in the Collaborative Cross.G3 (Bethesda). 2017 Jun 7;7(6):1665-1682. doi: 10.1534/g3.117.041624. G3 (Bethesda). 2017. PMID: 28592649 Free PMC article.
-
Influenza H3N2 infection of the collaborative cross founder strains reveals highly divergent host responses and identifies a unique phenotype in CAST/EiJ mice.BMC Genomics. 2016 Feb 27;17:143. doi: 10.1186/s12864-016-2483-y. BMC Genomics. 2016. PMID: 26921172 Free PMC article.
-
Host genetic variability and determinants of severe COVID-19.Trends Genet. 2023 Mar;39(3):169-171. doi: 10.1016/j.tig.2022.10.003. Epub 2022 Nov 12. Trends Genet. 2023. PMID: 36379742 Free PMC article.
-
Lymphocytic choriomeningitis virus Clone 13 infection causes either persistence or acute death dependent on IFN-1, cytotoxic T lymphocytes (CTLs), and host genetics.Proc Natl Acad Sci U S A. 2018 Aug 14;115(33):E7814-E7823. doi: 10.1073/pnas.1804674115. Epub 2018 Jul 30. Proc Natl Acad Sci U S A. 2018. PMID: 30061383 Free PMC article.
References
-
- Albright F.S., Orlando P., Pavia A.T., Jackson G.G., Cannon Albright L.A. Evidence for a heritable predisposition to death due to influenza. The Journal of Infectious Diseases. 2008;197(1):18–24. - PubMed
-
- Andrews T.D., Whittle B., Field M.A., Balakishnan B., Zhang Y., Shao Y. Massively parallel sequencing of the mouse exome to accurately identify rare, induced mutations: An immediate source for thousands of new mouse models. Open Biology. 2012;2(5):120061. doi: 10.1098/rsob.120061. - DOI - PMC - PubMed
-
- Bieber A.J., Suwansrinon K., Kerkvliet J., Zhang W., Pease L.R., Rodriguez M. Allelic variation in the Tyk2 and EGF genes as potential genetic determinants of CNS repair. Proceedings of the National Academy of Sciences of the United States of America. 2010;107(2):792–797. doi: 10.1073/pnas.0906589107. - DOI - PMC - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
