

Adaptation to background light enables contrast coding at rod bipolar cell synapses
- PMID: 24373883
- PMCID: PMC4267681
- DOI: 10.1016/j.neuron.2013.10.054
Adaptation to background light enables contrast coding at rod bipolar cell synapses
Abstract
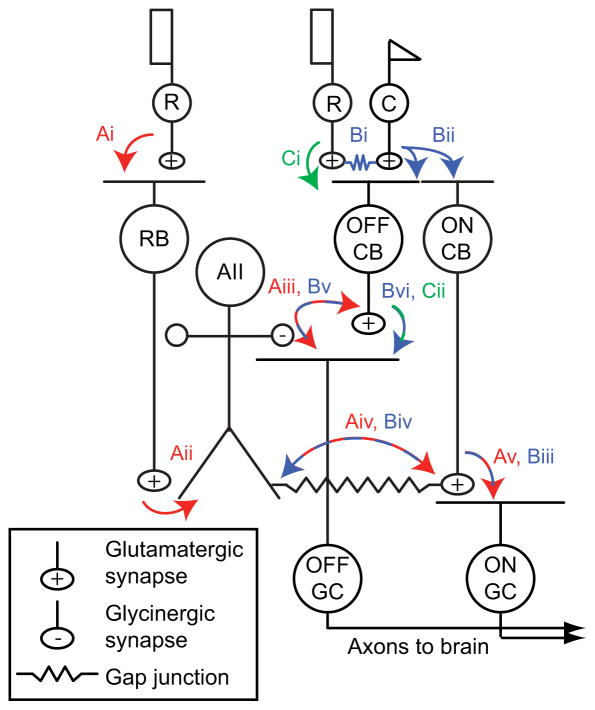
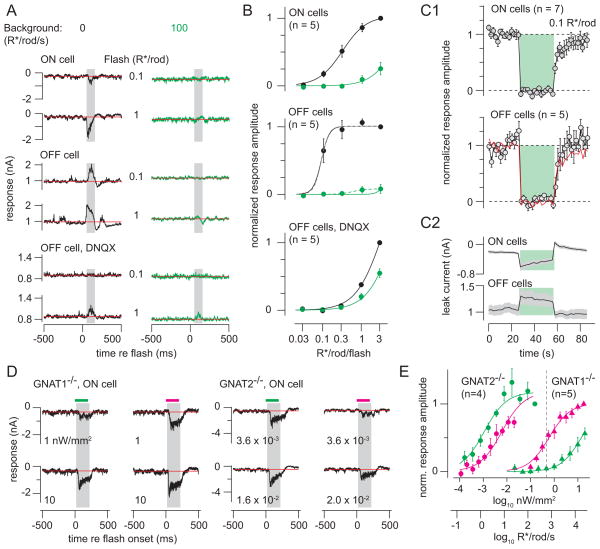
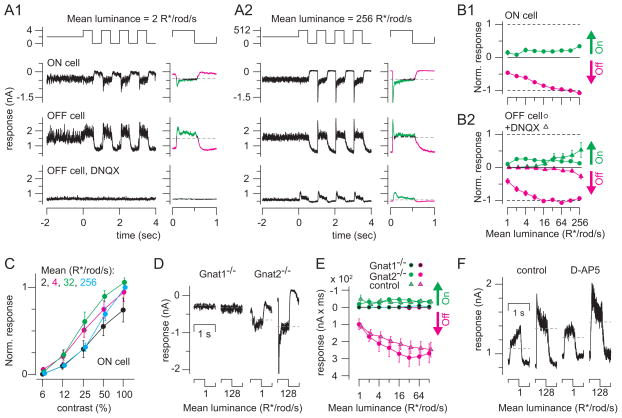
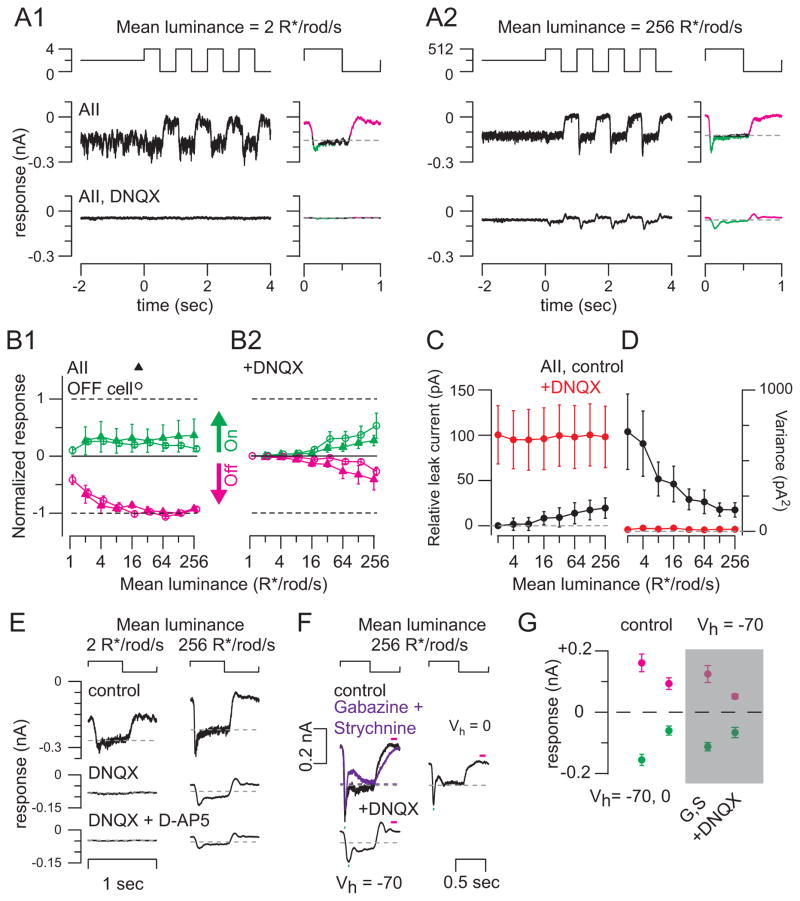
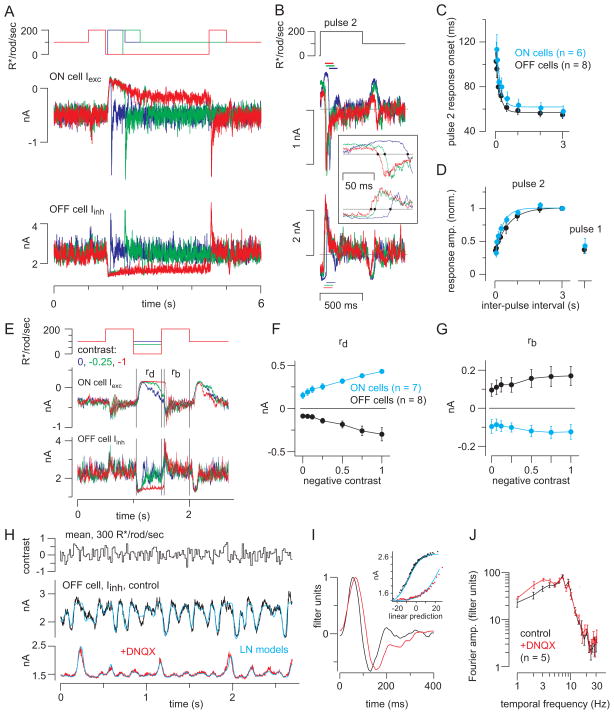
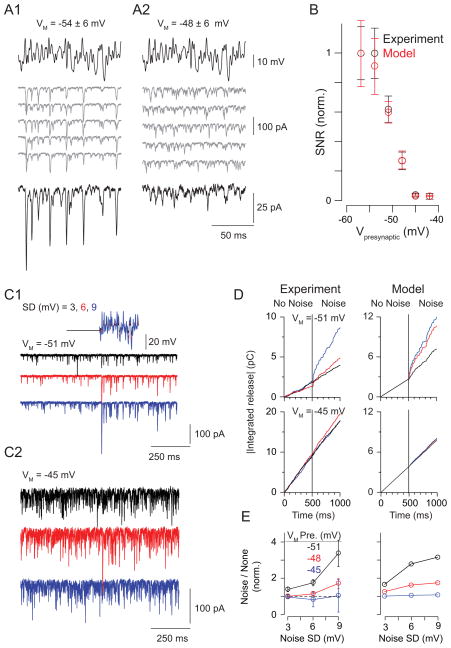
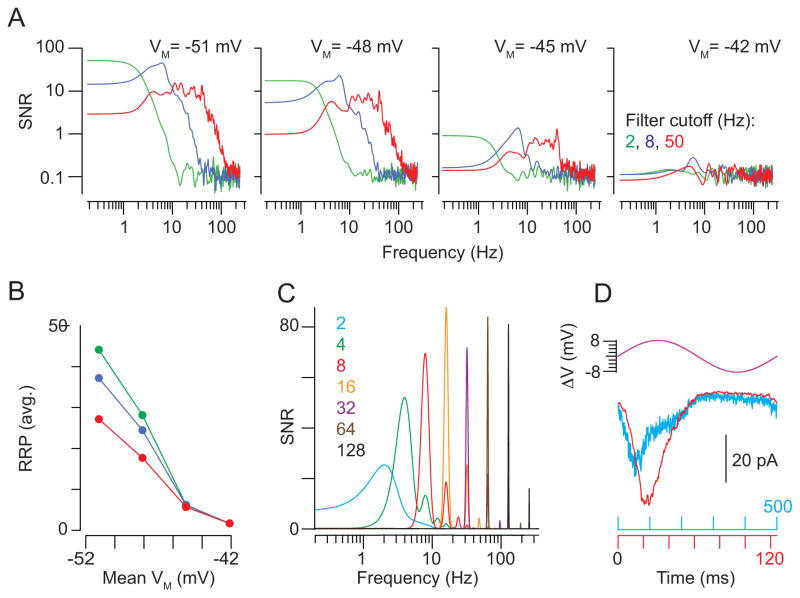
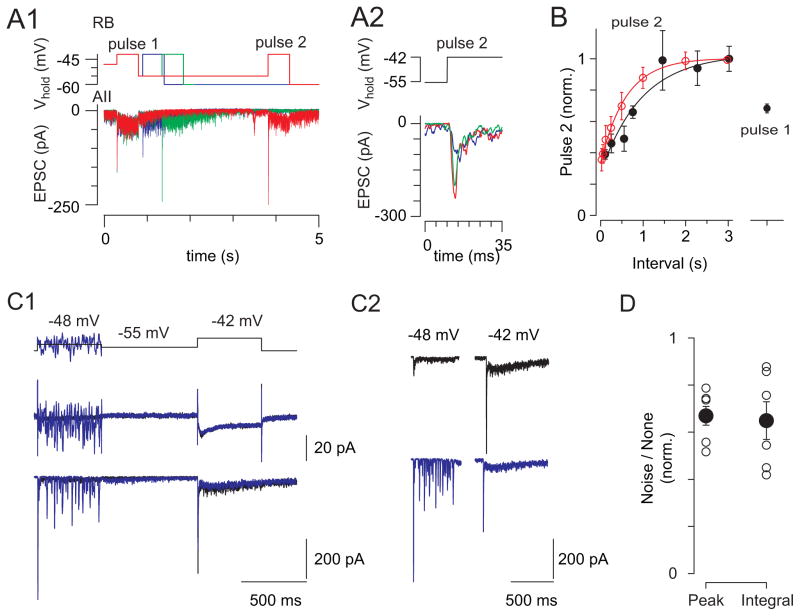
Rod photoreceptors contribute to vision over an ∼ 6-log-unit range of light intensities. The wide dynamic range of rod vision is thought to depend upon light intensity-dependent switching between two parallel pathways linking rods to ganglion cells: a rod → rod bipolar (RB) cell pathway that operates at dim backgrounds and a rod → cone → cone bipolar cell pathway that operates at brighter backgrounds. We evaluated this conventional model of rod vision by recording rod-mediated light responses from ganglion and AII amacrine cells and by recording RB-mediated synaptic currents from AII amacrine cells in mouse retina. Contrary to the conventional model, we found that the RB pathway functioned at backgrounds sufficient to activate the rod → cone pathway. As background light intensity increased, the RB's role changed from encoding the absorption of single photons to encoding contrast modulations around mean luminance. This transition is explained by the intrinsic dynamics of transmission from RB synapses.
Copyright © 2014 Elsevier Inc. All rights reserved.
Figures
References
-
- Applebury ML, Antoch MP, Baxter LC, Chun LL, Falk JD, Farhangfar F, Kage K, Krzystolik MG, Lyass LA, Robbins JT. The murine cone photoreceptor: a single cone type expresses both S and M opsins with retinal spatial patterning. Neuron. 2000;27:513–523. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
