

IpsA, a novel LacI-type regulator, is required for inositol-derived lipid formation in Corynebacteria and Mycobacteria
- PMID: 24377418
- PMCID: PMC3899939
- DOI: 10.1186/1741-7007-11-122
IpsA, a novel LacI-type regulator, is required for inositol-derived lipid formation in Corynebacteria and Mycobacteria
Abstract
Background: The development of new drugs against tuberculosis and diphtheria is focused on disrupting the biogenesis of the cell wall, the unique architecture of which confers resistance against current therapies. The enzymatic pathways involved in the synthesis of the cell wall by these pathogens are well understood, but the underlying regulatory mechanisms are largely unknown.
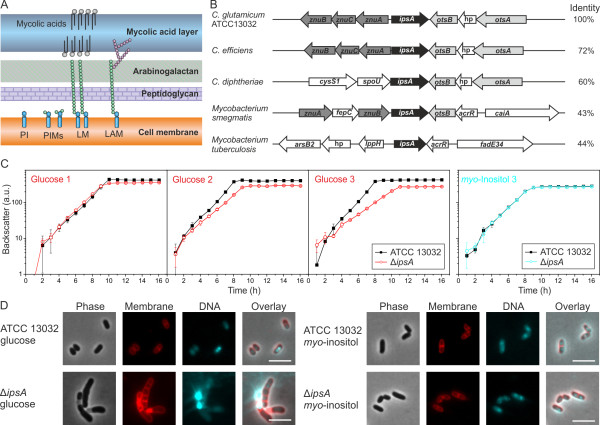
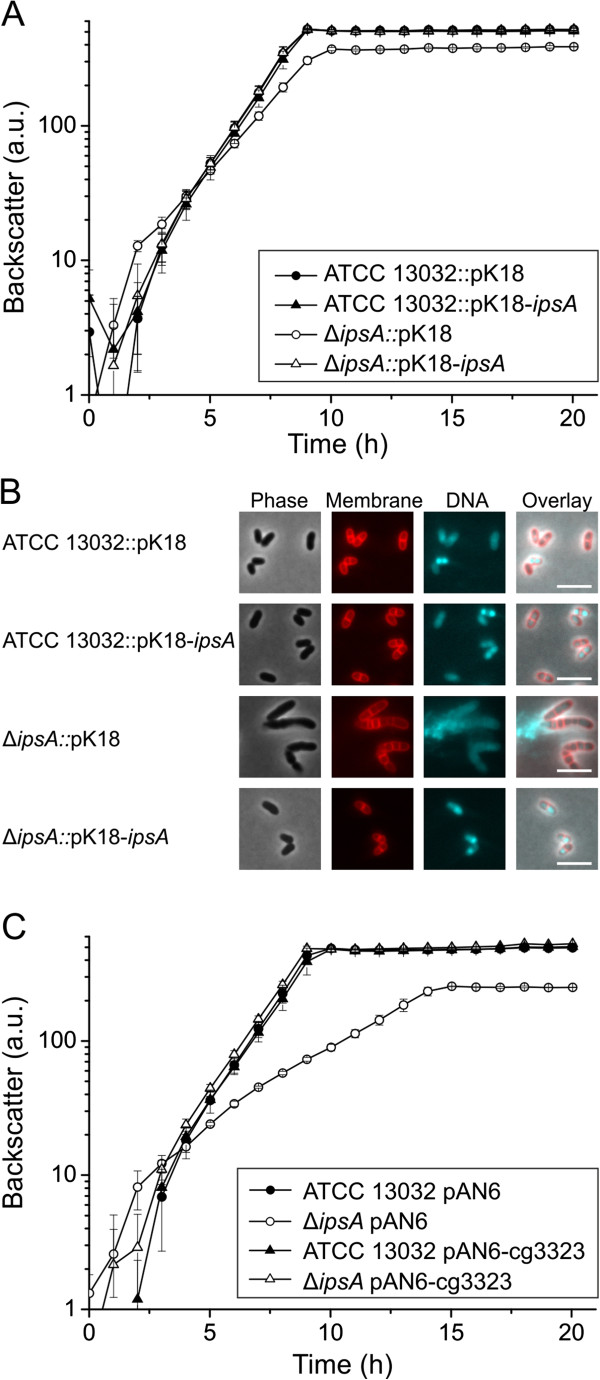
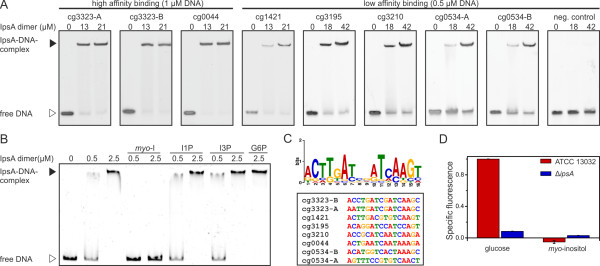
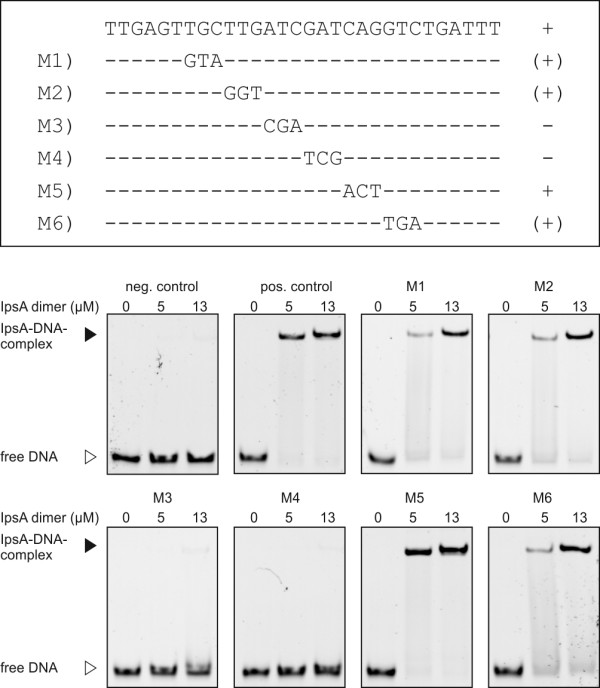
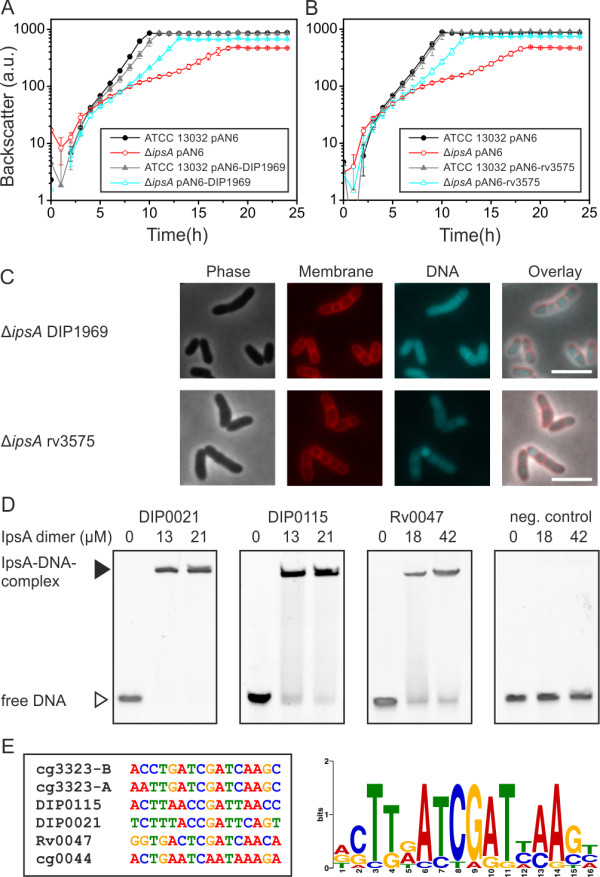
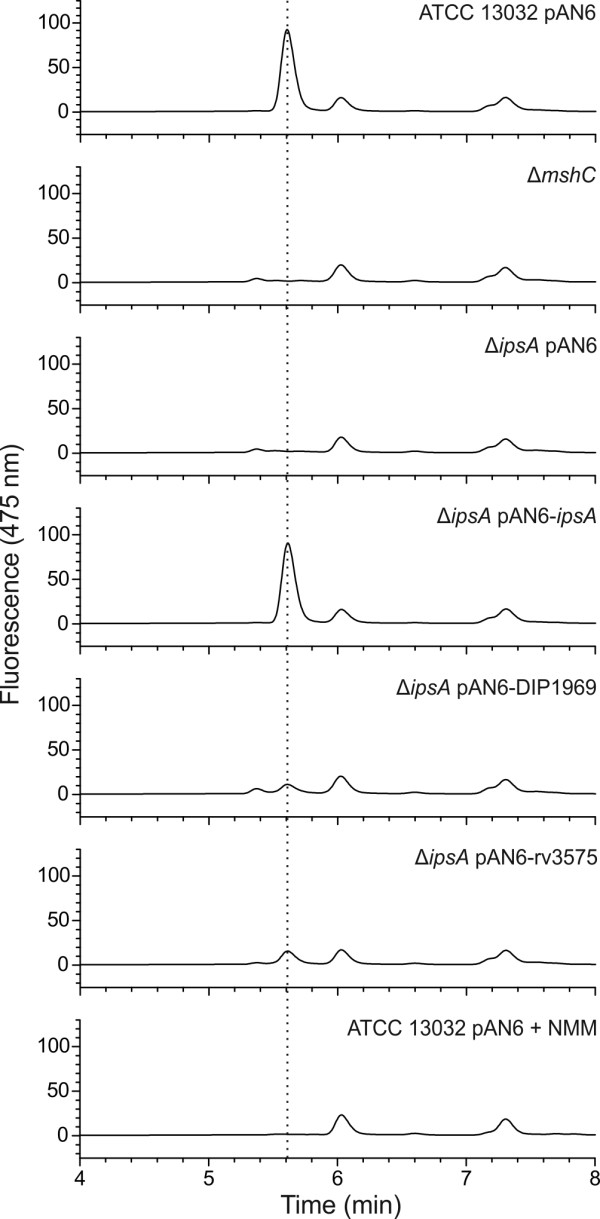
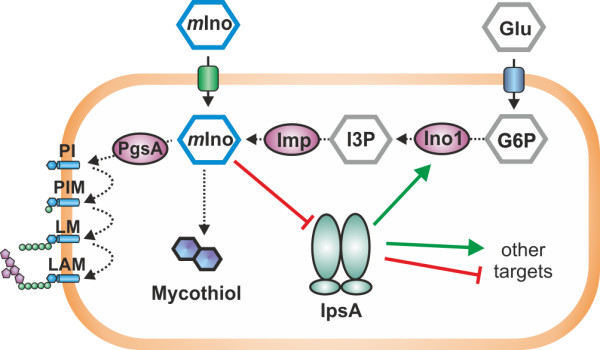
Results: Here, we characterize IpsA, a LacI-type transcriptional regulator conserved among Mycobacteria and Corynebacteria that plays a role in the regulation of cell wall biogenesis. IpsA triggers myo-inositol formation by activating ino1, which encodes inositol phosphate synthase. An ipsA deletion mutant of Corynebacterium glutamicum cultured on glucose displayed significantly impaired growth and presented an elongated cell morphology. Further studies revealed the absence of inositol-derived lipids in the cell wall and a complete loss of mycothiol biosynthesis. The phenotype of the C. glutamicum ipsA deletion mutant was complemented to different extend by homologs from Corynebacterium diphtheriae (dip1969) and Mycobacterium tuberculosis (rv3575), indicating the conserved function of IpsA in the pathogenic species. Additional targets of IpsA with putative functions in cell wall biogenesis were identified and IpsA was shown to bind to a conserved palindromic motif within the corresponding promoter regions. Myo-inositol was identified as an effector of IpsA, causing the dissociation of the IpsA-DNA complex in vitro.
Conclusions: This characterization of IpsA function and of its regulon sheds light on the complex transcriptional control of cell wall biogenesis in the mycolata taxon and generates novel targets for drug development.
Figures

References
-
- Eggeling L, Besra GS, Alderwick L. In: Corynebacteria: genomics and molecular biology. Burkovski A, editor. Wymondham, UK: Caister Academic Press; 2008. Structure and synthesis of the cell wall; pp. 267–294.
-
- Fahey RC. Glutathione analogs in prokaryotes. Biochim Biophys Acta. 1830;11:3182–3198. - PubMed
-
- Morita YS, Fukuda T, Sena CBC, Yamaryo-Botte Y, McConville MJ, Kinoshita T. Inositol lipid metabolism in mycobacteria: biosynthesis and regulatory mechanisms. Biochim Biophys Acta. 1810;11:630–641. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
