

Identification of a crucial residue required for Staphylococcus aureus LukAB cytotoxicity and receptor recognition
- PMID: 24379286
- PMCID: PMC3958006
- DOI: 10.1128/IAI.01444-13
Identification of a crucial residue required for Staphylococcus aureus LukAB cytotoxicity and receptor recognition
Abstract
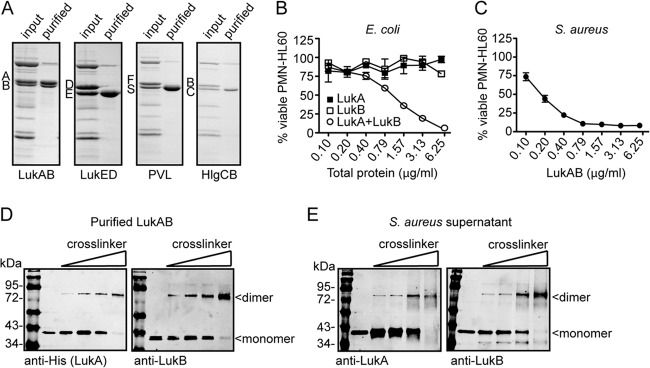
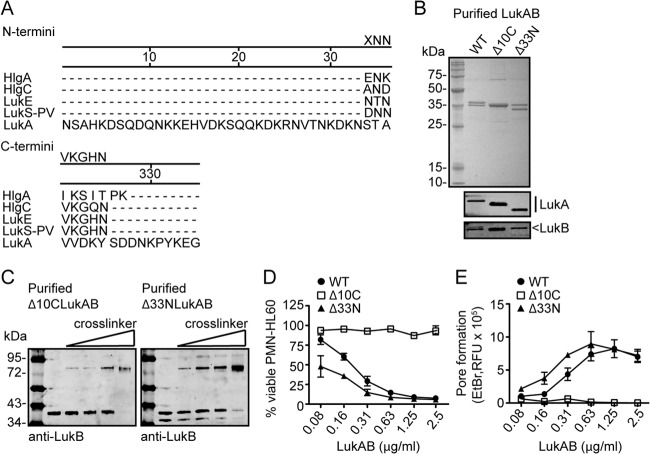
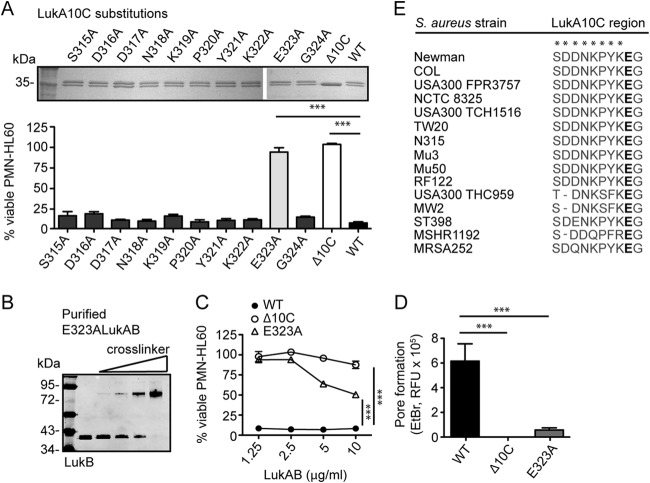
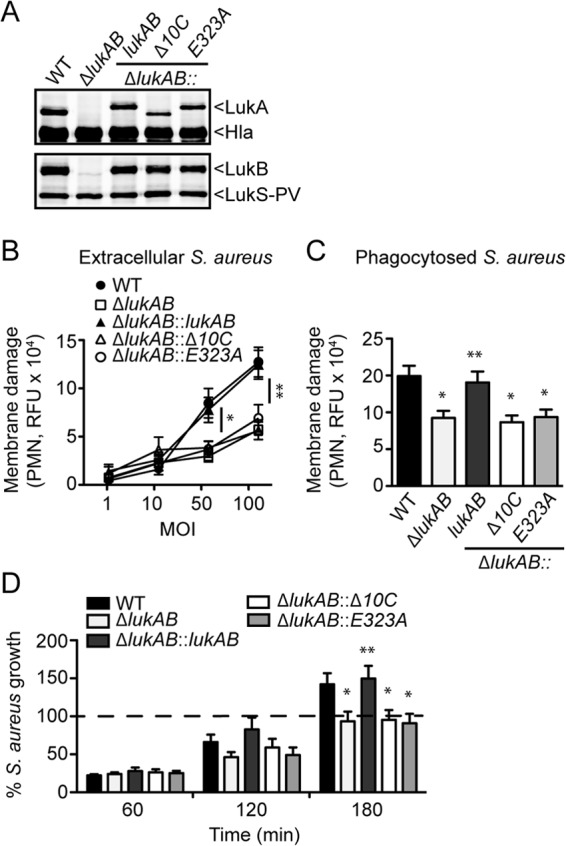
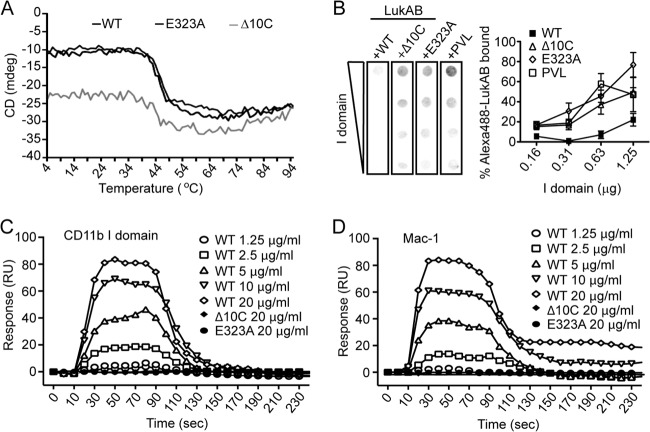
The bicomponent leukotoxins produced by Staphylococcus aureus kill host immune cells through osmotic lysis by forming β-barrel pores in the host plasma membrane. The current model for bicomponent pore formation proposes that octameric pores, comprised of two separate secreted polypeptides (S and F subunits), are assembled from water-soluble monomers in the extracellular milieu and multimerize on target cell membranes. However, it has yet to be determined if all staphylococcal bicomponent leukotoxin family members exhibit these properties. In this study, we report that leukocidin A/B (LukAB), the most divergent member of the leukotoxin family, exists as a heterodimer in solution rather than two separate monomeric subunits. Notably, this property was found to be associated with enhanced toxin activity. LukAB also differs from the other bicomponent leukotoxins in that the S subunit (LukA) contains 33- and 10-amino-acid extensions at the N and C termini, respectively. Truncation mutagenesis revealed that deletion of the N terminus resulted in a modest increase in LukAB cytotoxicity, whereas the deletion of the C terminus rendered the toxin inactive. Within the C terminus of LukA, we identified a glutamic acid at position 323 that is critical for LukAB cytotoxicity. Furthermore, we discovered that this residue is conserved and required for the interaction between LukAB and its cellular target CD11b. Altogether, these findings provide an in-depth analysis of how LukAB targets neutrophils and identify novel targets suitable for the rational design of anti-LukAB inhibitors.
Figures
Similar articles
-
Unlatching of the stem domains in the Staphylococcus aureus pore-forming leukocidin LukAB influences toxin oligomerization.J Biol Chem. 2023 Dec;299(12):105321. doi: 10.1016/j.jbc.2023.105321. Epub 2023 Oct 4. J Biol Chem. 2023. PMID: 37802313 Free PMC article.
-
Staphylococcus aureus LukAB cytotoxin kills human neutrophils by targeting the CD11b subunit of the integrin Mac-1.Proc Natl Acad Sci U S A. 2013 Jun 25;110(26):10794-9. doi: 10.1073/pnas.1305121110. Epub 2013 Jun 10. Proc Natl Acad Sci U S A. 2013. PMID: 23754403 Free PMC article.
-
Genetic variation of staphylococcal LukAB toxin determines receptor tropism.Nat Microbiol. 2021 Jun;6(6):731-745. doi: 10.1038/s41564-021-00890-3. Epub 2021 Apr 19. Nat Microbiol. 2021. PMID: 33875847 Free PMC article.
-
The bicomponent pore-forming leucocidins of Staphylococcus aureus.Microbiol Mol Biol Rev. 2014 Jun;78(2):199-230. doi: 10.1128/MMBR.00055-13. Microbiol Mol Biol Rev. 2014. PMID: 24847020 Free PMC article. Review.
-
Staphylococcus aureus pore-forming toxins: The interface of pathogen and host complexity.Semin Cell Dev Biol. 2017 Dec;72:101-116. doi: 10.1016/j.semcdb.2017.04.003. Epub 2017 Apr 23. Semin Cell Dev Biol. 2017. PMID: 28445785 Free PMC article. Review.
Cited by
-
Vaccination With Detoxified Leukocidin AB Reduces Bacterial Load in a Staphylococcus aureus Minipig Deep Surgical Wound Infection Model.J Infect Dis. 2022 Apr 19;225(8):1460-1470. doi: 10.1093/infdis/jiab219. J Infect Dis. 2022. PMID: 33895843 Free PMC article.
-
Unbiased Identification of Immunogenic Staphylococcus aureus Leukotoxin B-Cell Epitopes.Infect Immun. 2020 Mar 23;88(4):e00785-19. doi: 10.1128/IAI.00785-19. Print 2020 Mar 23. Infect Immun. 2020. PMID: 32014894 Free PMC article.
-
Staphylococcus aureus Leukocidin LukED and HIV-1 gp120 Target Different Sequence Determinants on CCR5.mBio. 2016 Dec 13;7(6):e02024-16. doi: 10.1128/mBio.02024-16. mBio. 2016. PMID: 27965453 Free PMC article.
-
Host-Pathogen Interface: Progress in Understanding the Pathogenesis of Infection Due to Multidrug-Resistant Bacteria in the Intensive Care Unit.J Infect Dis. 2017 Feb 15;215(suppl_1):S1-S8. doi: 10.1093/infdis/jiw405. J Infect Dis. 2017. PMID: 28375516 Free PMC article.
-
Residues essential for Panton-Valentine leukocidin S component binding to its cell receptor suggest both plasticity and adaptability in its interaction surface.PLoS One. 2014 Mar 18;9(3):e92094. doi: 10.1371/journal.pone.0092094. eCollection 2014. PLoS One. 2014. PMID: 24643034 Free PMC article.
References
-
- Panton PN, Valentine FCO. 1932. Staphylococcal toxin. Lancet i:506–508
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
