

Review
doi: 10.1152/physiol.00035.2013.
Respiratory rhythm generation in vivo
Affiliations
- PMID: 24382872
- PMCID: PMC3929116
- DOI: 10.1152/physiol.00035.2013
Item in Clipboard
Review
Respiratory rhythm generation in vivo
Physiology (Bethesda).
2014 Jan.
Abstract
The cellular and circuit mechanisms generating the rhythm of breathing in mammals have been under intense investigation for decades. Here, we try to integrate the key discoveries into an updated description of the basic neural processes generating respiratory rhythm under in vivo conditions.
Conflict of interest statement
No conflicts of interest, financial or otherwise, are declared by the author(s).
Figures
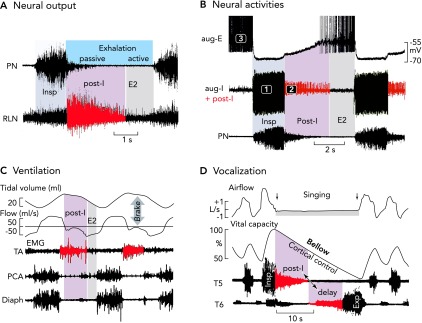
Neural and mechanical phases of a respiratory cycle in vivo A: phrenic and recurrent laryngeal (RLN) nerve recordings in the anesthetized cat and rat reveal an augmenting inspiratory burst that ends abruptly. Thereafter, there appears a declining post-inspiratory discharge particularly strong in the RLN representing the neural control of laryngeal adductor muscles (83). B: simultaneous in vivo triplet recordings from three characteristic types of neuron in the anesthetized cat reveal a regular sequence of inspiratory (phase 1), post-inspiratory (phase 2), and late-expiratory (phase 3) discharges (action potentials are truncated) (77). C: the post-inspiratory discharge of laryngeal nerves activates a mechanical brake of the expiratory airflow and a holding of lung volume. Recordings are from the thyroarytenoid constrictor muscle (TA), the posterior cricoarythyoid dilator muscle (PCA), and the inspiratory diaphragm (Diaph) of halothane anesthetized lamb (see modified Figure 5 from Ref. 28). D: post-inspiratory activity controls vocalization. Recordings were from human inspiratory intercostal muscles in T5 and expiratory muscles in T6, and clearly show a significance of the post-inspiratory control of inspiratory and expiratory muscle activity during singing, which requires tight control of the breathing rhythm (85).
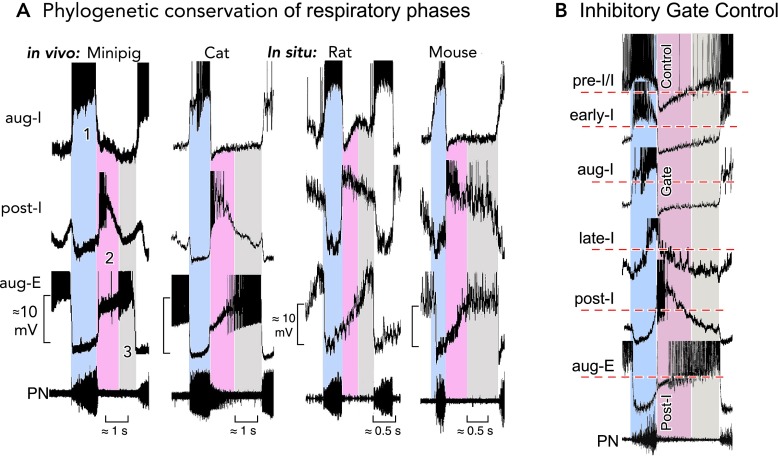
Characteristic neuronal activities revealing a conserved respiratory network operation A: the three-phased organization of respiratory activities in the ventrolateral medulla containing the pre-BötC and BötC regions is highly conserved in mammalian species. Characteristic patterns of neuronal discharges during each phase (1–3) and the underlying temporal fluctuations of membrane potentials revealed by intracellular recordings are very similar in mini-pig (41) and cat (72) in vivo, as well as in rat and mouse brain stem-spinal cord preparations in situ (9). Recordings are magnified in each panel to emphasize underlying membrane potential trajectories. Action potentials have been truncated. B: aligning the activities (truncated action potentials) of different types of respiratory neurons in the pre-BötC and BötC (from the cat in this illustration) to the rhythm defined by the output phrenic nerve discharges identifies post-I inhibition that produces dominant inhibitory synaptic volleys in pre-I/I, early-I, aug-I, and aug-E/E2 neurons and, therefore, exerts an effective control of rhythm generation by activity and voltage resetting, as originally described by Richter (75).
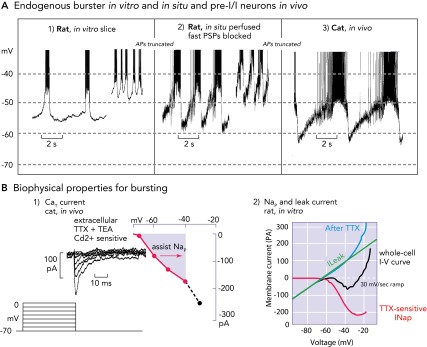
Biophysical properties and bursting behavior of pre-BötC inspiratory neurons A: subpopulations of rodent pre-BötC neurons exhibit voltage-dependent endogenous rhythmic bursting in vitro (A1) after blockade of fast excitatory synaptic transmission (adapted from Ref. and used with permission) or in situ (A2) after blockade of fast synaptic transmission (93). The membrane potentials for onset of rhythmic bursting in all cases is −60 mV, and the bursting frequency increases progressively with depolarization up to a baseline at approximately −45 mV. Identified eBNs reveal similar interburst voltage trajectories as recorded from pre-I/I neurons in vivo (A3), some of which may have intrinsic bursting behavior (58, 64). Recorded in vivo, these neurons also start to depolarize for bursting at a voltage range of approximately −60 mV. As synaptic interactions are intact in vivo, the membrane potential trajectory is controlled by postsynaptic inhibition during the post-inspiratory phase, during which endogenous bursting is suppressed. All action potentials are truncated. B1: when conditioned with hyperpolarizing pre-pulses, all groups of respiratory neurons generate a CaT current already at negative voltages of −80 mV that can depolarize neurons to spike activation threshold (64). B2: the underlying biophysical properties for in vitro and in situ endogenous bursting are primarily a TTX-sensitive persistent Na+ channel (Nap) and an ohmic-like Kleak conductance (34). The Nap current-voltage (I-V) relation (red curve), obtained from a slow voltage clamp ramp protocol applied to in vitro eBNs, is revealed after blocking the current with TTX and subtracting the resultant I-V relation from the whole cell I-V curve measured before TTX application. The K+-dominated leak I-V relation (green line) is obtained from the linear region of the whole-cell I-V curve (after Ref. 34).
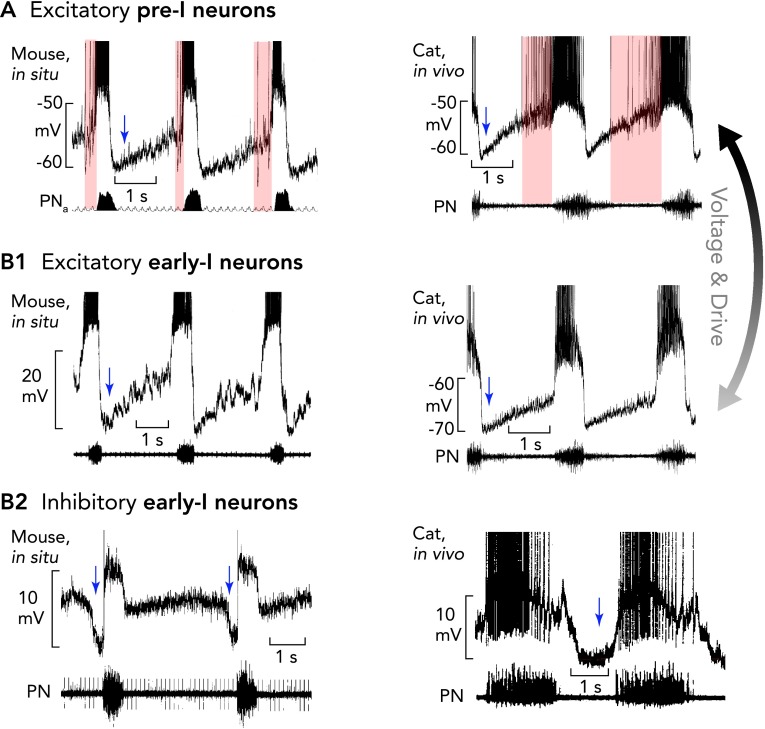
Pre-I/I and two types of early-I neurons The pre-BötC contains excitatory pre-I/I neurons (A) and two types of early-I spiking neurons [excitatory (B1) and inhibitory (B2)], which are considered to be important for inspiratory phase generation (pre-I/I and early-I excitatory neurons) and coordinating the inspiratory-expiratory phase transition (early-I inhibitory neurons). Recordings in mice are shown in at left and recordings in anesthetized cats at right. All action potentials are truncated. A and B1: characteristic of the pre-I/I neurons (onset and duration of their variable pre-I firing is indicated by pink areas) and early-I excitatory neurons is a lack of, or only weak, synaptic inhibition during stage 2 expiration. When hyperpolarized, pre-I/I neurons acquire an activity pattern similar to early-I excitatory neurons. The membrane potential trajectories and rapid onset of spiking of early-I excitatory neurons, which are thought to be part of the heterogenous excitatory neuron population critical for inspiratory phase generation, probably result from lower tonic excitation that regulate their busting behavior in the intact network. B2: inhibitory early-I neurons receive strong inhibition during the E2 phase (blue shaded region) and are important components of the inhibitory connectome (see FIGURE 6) controlling generation of expiratory phase activity. Periods of postsynaptic inhibition are indicated by blue arrows.
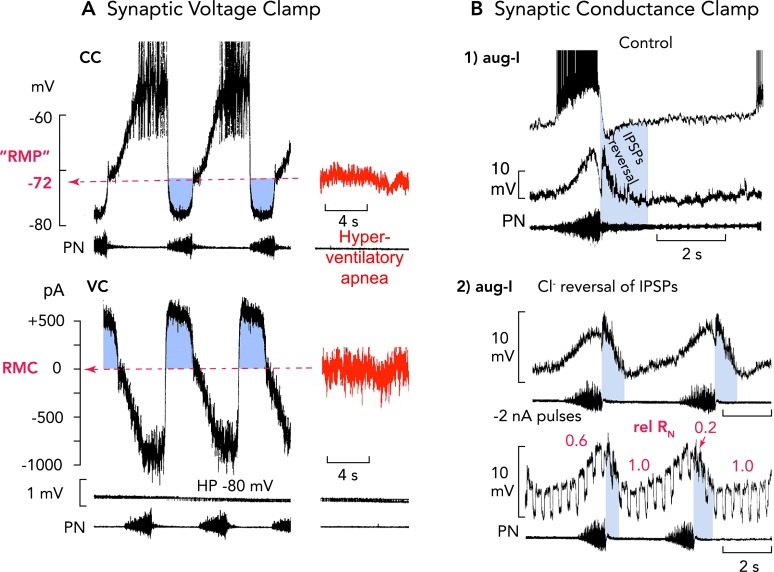
“Restless” respiratory neurons: a bombardment of excitatory and inhibitory synaptic inputs controls ongoing membrane potential oscillations A: high-frequency alternating single-electrode current (CC) and voltage-clamp (VC) recordings [top and bottom, respectively; holding potential (HP)] from an expiratory neuron, as an example (76), reveals that neurons operating in the active respiratory network do not show any resting membrane potential. There are ongoing oscillations of membrane potential generated by alternating excitatory and inhibitory (blue regions) synaptic volleys. A fictive resting membrane potential (RMP) where the net current is 0 pA (RMC, bottom) at about −70 mV could only be estimated when animals were hyperventilated to provoke central apnea. Characteristic for inspiratory and expiratory neurons are very effective volleys of inhibitory currents causing a functional voltage clamp of neurons close to the chloride equilibrium potential (ECl) at approximately −80 mV. Action potentials are truncated. B: the prominent inhibitory synaptic volleys also produce a significant fall of neuronal input resistance (RN), which exerts a conductance clamp, because it effectively shunts endogenous and exogenous synaptic currents (72, 74). In all inspiratory neurons (examples of aug-I neurons shown), the most obvious RN fall by up to 80% occurs at the onset of post-I inhibition, as identified by IPSPs polarity reversal (blue regions) after intracellular chloride injection (71). Action potentials are truncated.
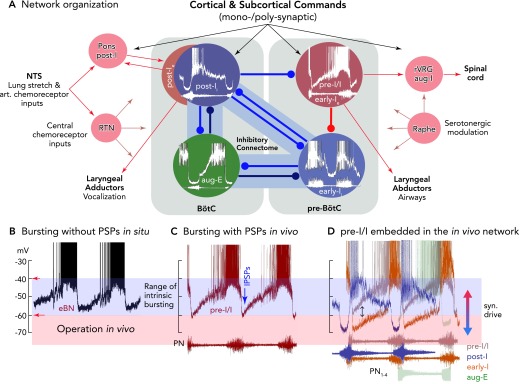
Functional model of the rhythmogenic respiratory network organization, including control by afferent inputs and output functions of neurons A: the different patterns of synaptic inputs to the neurons of the network indicate that antagonistic (reciprocal) inhibition is controlling in vivo rhythm generation and coordinating inspiratory and expiratory activity phases, as well as operating under control of various input signals. There are various sorts of antagonism organized by inhibitory connectomes, as proposed by the authors (, , 92) and supported by computational simulations (e.g., Refs. 58, 80, 89) as a “proof of principle.” The most important antagonistic connectivity involved in rhythm generation appears to exist between early-I and post-I inhibitory neurons and their connections with pre-I/I excitatory neurons, allowing inspiratory-post-inspiratory activity coordination, besides other connectomes of aug-E neurons to early-I and post-I neurons. Inhibitory connections are depicted as blue lines, excitatory connections as red lines, and other afferent inputs as gray lines. B: bursting patterns of eBNs recorded in situ, as revealed after block of synaptic transmission (without PSPs), illustrating membrane potential trajectories and approximate voltage range for intrinsic rhythmic bursting (see also FIGURE 4A). C: pre-I/I neurons/eBNs (red trace) embedded in the core circuitry in vivo and in situ and presumably receiving tonic excitation have depolarizing membrane potential trajectories and generate bursts (see FIGURE 4A) between voltages of approximately −60 to −40 mV, where the membrane potential trajectory is effectively controlled by post-I inhibition (IPSPs). D: using the onset and end of phrenic nerve bursts as a relative time coordinate shows the sequences of neuronal activities and membrane potential trajectories of pre-I/I, post-I, early-I, and aug-E neurons of the connectome shown in A. Potential pre-I/I neurons with stronger post-I and late-expiratory inhibition or receiving lower tonic excitation will behave as early-I bursting neurons (see color code for neuron types). All action potentials are truncated.
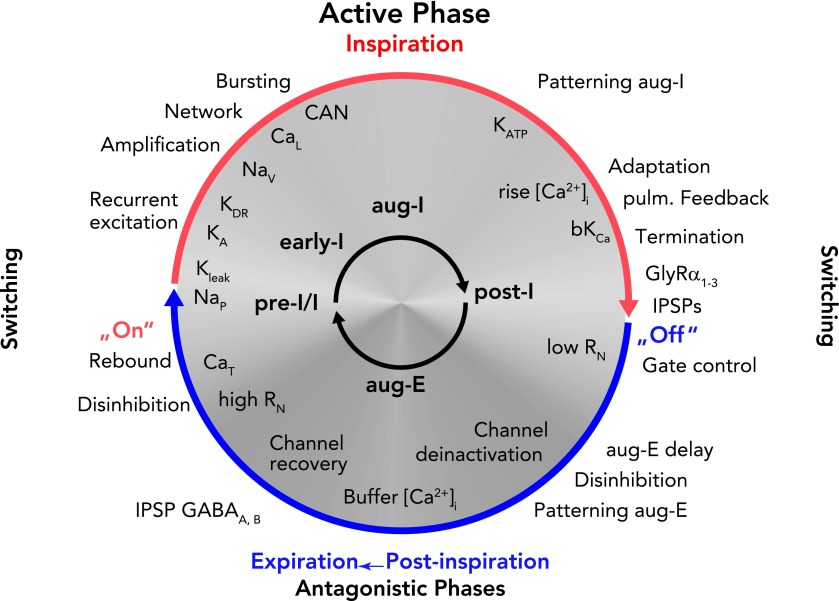
Schematic representation of the dynamic interplay of synaptic processes and neuronal biophysical properties determining rhythm generation and neuronal activity patterns during a respiratory cycle The ongoing rhythmic cycling of respiratory phases results from a coalition between synaptic inputs to neurons and intrinsic neuronal biophysical properties (65). Network-related synaptic events [recurrent excitation, and inhibitory glycinergic (GlyRa1–3) or GABAergic (GABAA,B) neurotransmitter-related inhibition (IPSPs) or disinhibition] are indicated outside the circle. Note that there is a continuous inflow of synaptic activity from pulmonary and chemoreceptive afferents, as well as pontine and medullary reticular formation inputs. Intrinsic neuronal biophysical processes, including membrane channel activation, recovery from inactivation, deinactivation under membrane voltage control, as well as activity-related Ca2+-dependent processes, are indicated inside the circle in white lettering. Synaptic processes in the rhythmically active respiratory circuits control membrane potentials in neurons in the subthreshold voltage range to orchestrate expression of intrinsic membrane conductances, including burst-promoting conductances (CaT and NaP) as well as K+ conductances (KLeak, KA) that regulate onset of bursting. During neuronal spiking activity, Na+ and Ca2+ fluxes together with CAN augment neuronal activities to generate the active phases of the cycle. Ca2+ fluxes also activate bK(Ca) to induce spike-frequency adaptation that, in addition to synaptic inhibition, contributes to termination of active phases. The figure should be read clockwise. For further explanation, see text.
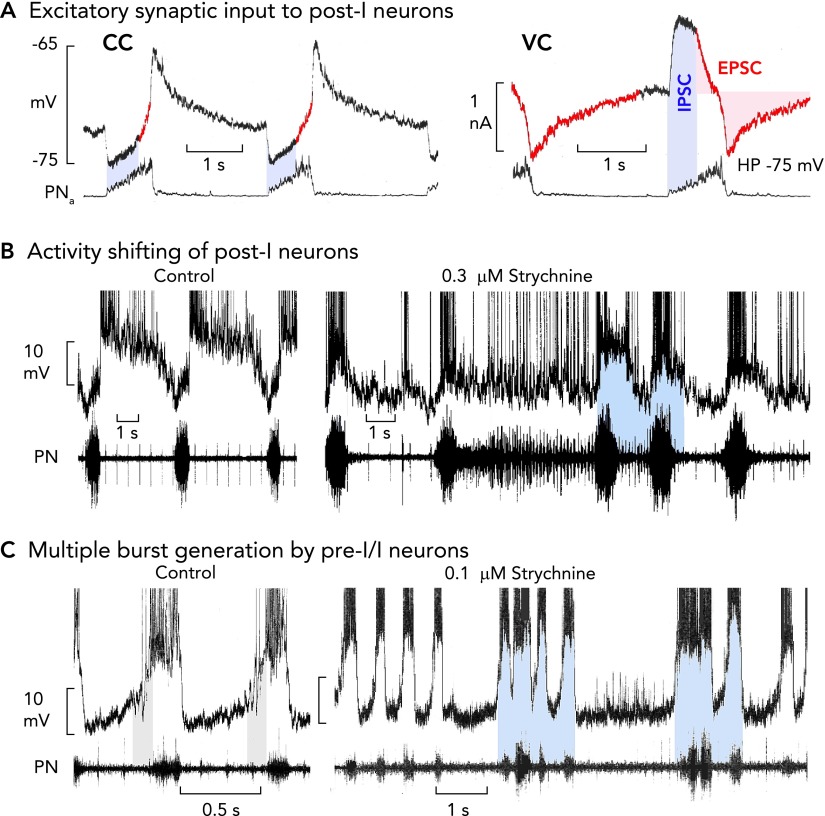
Perturbations of medullary respiratory neuron and network activities during failure of glycinergic inhibition A: post-I neurons receive an excitatory synaptic drive already during inspiration, shown in current clamp recording (CC). This becomes clear when neurons are voltage clamped [VC; holding potential (HP)] at normal voltages, revealing onset of excitatory inward currents during inspiration (see red colored traces; also see Refs. 47, 76). B: blockade of glycinergic inhibition provokes a shifting of post-I discharges into the period of inspiration due to the inspiratory excitatory synaptic input (9). The consequence is a recurring interruption of regular bursting leading to inspiratory and post-inspiratory doublet or multiple bursts, and persisting inspiratory (apneustic) activity is transmitted to phrenic nerve output (11). C: strychnine augments inspiratory burst amplitudes and discharge frequency of pre-I/I neurons due to the block of the normal glycine receptor-mediated steep membrane hyperpolarization after the burst. The activity shifting of post-I discharge into inspiration provokes a dynamic struggle between post-I neurons and their counterparts in the connectome, with pre-I/I bursters trying to start an inspiration against the shifted post-I neuron post-I inhibition that is reduced, but not completely blocked, by strychnine (11). All action potentials in B and C are truncated.
References
-
- Alheid GF, Milsom WK, McCrimmon DR. Pontine influences on breathing: an overview. Respir Physiol Neurobiol 143: 105–114, 2004 - PubMed
-
- Backman SB, Anders C, Ballantyne D, Röhrig N, Camerer H, Mifflin S, Jordan D, Dickhaus H, Spyer KM, Richter DW. Evidence for a monosynaptic connection between slowly adapting pulmonary stretch receptor afferents and inspiratory beta neurones. Pflügers Arch 402: 129–136, 1984 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources