

The TRPM6 kinase domain determines the Mg·ATP sensitivity of TRPM7/M6 heteromeric ion channels
- PMID: 24385424
- PMCID: PMC3931078
- DOI: 10.1074/jbc.M113.512285
The TRPM6 kinase domain determines the Mg·ATP sensitivity of TRPM7/M6 heteromeric ion channels
Abstract
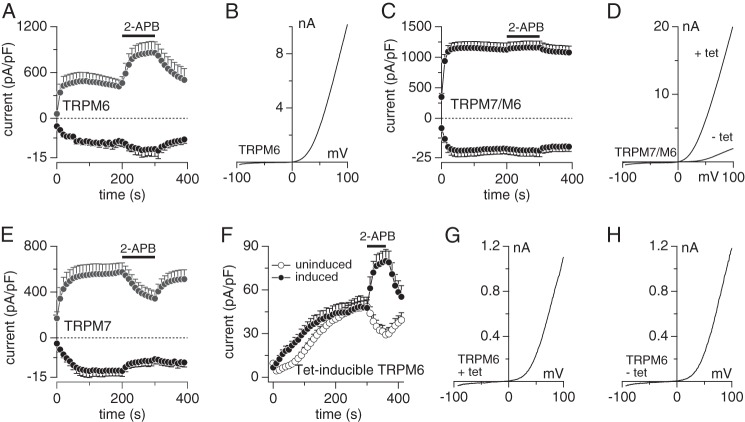
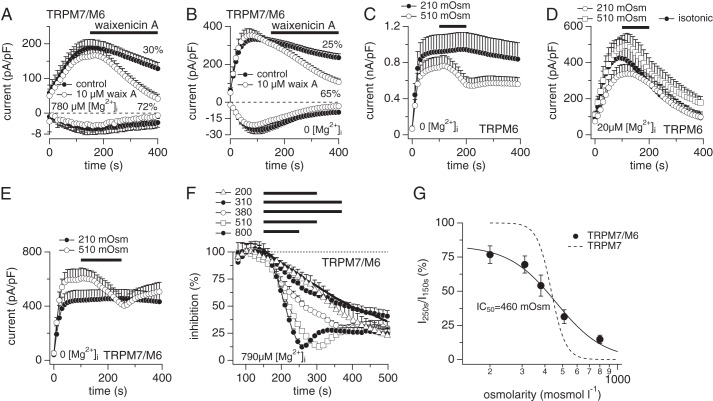
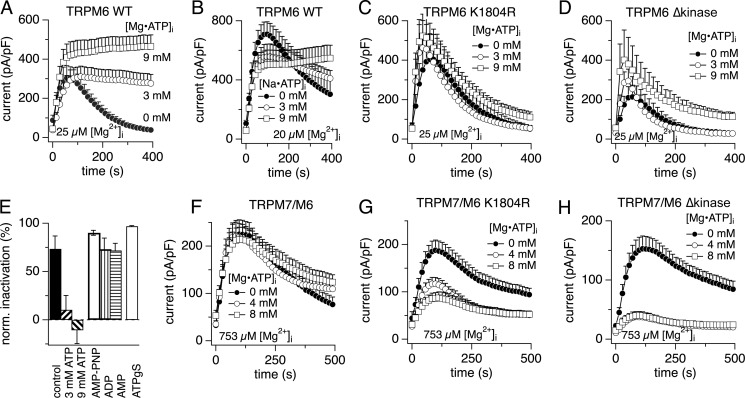
The transient receptor potential melastatin member 7 (TRPM7) and member 6 (TRPM6) are divalent cation channel kinases essential for magnesium (Mg(2+)) homeostasis in vertebrates. It remains unclear how TRPM6 affects divalent cation transport and whether this involves functional homomeric TRPM6 plasma membrane channels or heteromeric channel assemblies with TRPM7. We show that homomeric TRPM6 is highly sensitive to intracellular free Mg(2+) and therefore unlikely to be active at physiological levels of [Mg(2+)]i. Co-expression of TRPM7 and TRPM6 produces heteromeric TRPM7/M6 channels with altered pharmacology and sensitivity to intracellular Mg·ATP compared with homomeric TRPM7. Strikingly, the activity of heteromeric TRPM7/M6 channels is independent of intracellular Mg·ATP concentrations, essentially uncoupling channel activity from cellular energy status. Disruption of TRPM6 kinase phosphorylation activity re-introduces Mg·ATP sensitivity to the heteromeric channel similar to that of TRPM7. Thus, TRPM6 modulates the functionality of TRPM7, and the TRPM6 kinase plays a critical role in tuning the phenotype of the TRPM7·M6 channel complex.
Keywords: Magnesium; Metabolic Regulation; Protein Complexes; Protein Kinases; TRP channels.
Figures


References
-
- Walder R. Y., Landau D., Meyer P., Shalev H., Tsolia M., Borochowitz Z., Boettger M. B., Beck G. E., Englehardt R. K., Carmi R., Sheffield V. C. (2002) Mutation of TRPM6 causes familial hypomagnesemia with secondary hypocalcemia. Nat. Genet. 31, 171–174 - PubMed
-
- Schlingmann K. P., Weber S., Peters M., Niemann Nejsum L., Vitzthum H., Klingel K., Kratz M., Haddad E., Ristoff E., Dinour D., Syrrou M., Nielsen S., Sassen M., Waldegger S., Seyberth H. W., Konrad M. (2002) Hypomagnesemia with secondary hypocalcemia is caused by mutations in TRPM6, a new member of the TRPM gene family. Nat. Genet. 31, 166–170 - PubMed
-
- Nadler M. J., Hermosura M. C., Inabe K., Perraud A. L., Zhu Q., Stokes A. J., Kurosaki T., Kinet J. P., Penner R., Scharenberg A. M., Fleig A. (2001) LTRPC7 is a Mg·ATP-regulated divalent cation channel required for cell viability. Nature 411, 590–595 - PubMed
-
- Runnels L. W., Yue L., Clapham D. E. (2001) TRP-PLIK, a bifunctional protein with kinase and ion channel activities. Science 291, 1043–1047 - PubMed
-
- Voets T., Nilius B., Hoefs S., van der Kemp A. W., Droogmans G., Bindels R. J., Hoenderop J. G. (2004) TRPM6 forms the Mg2+ influx channel involved in intestinal and renal Mg2+ absorption. J. Biol. Chem. 279, 19–25 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
