

Review
doi: 10.1074/jbc.X114.547323.
Epub 2014 Jan 2.
The chemistry of regulation of genes and other things
Affiliations
- PMID: 24385432
- PMCID: PMC3937619
- DOI: 10.1074/jbc.X114.547323
Item in Clipboard
Review
The chemistry of regulation of genes and other things
J Biol Chem.
.
No abstract available
Keywords: Activators; Cooperativity; DNA-binding Protein; Gene Regulation; Genetic Switch; Nucleosome; Recruitment; Repressors; Specificity; Transcription.
Figures
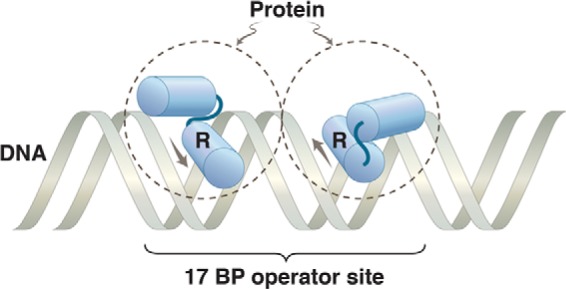
Protein dimer bound to DNA. The dashed circles represent two identical subunits of a protein bound to a 2-fold rotationally symmetric operator. The HTH motif on each monomer is indicated, with the recognition helix labeled R. The arrows show the direction N → C of the recognition helix in each monomer. We are dealing here with a simple binding reaction, so the protein rapidly comes off and rebinds. The image represents, in effect, a snapshot of the protein and DNA at an instant of binding.
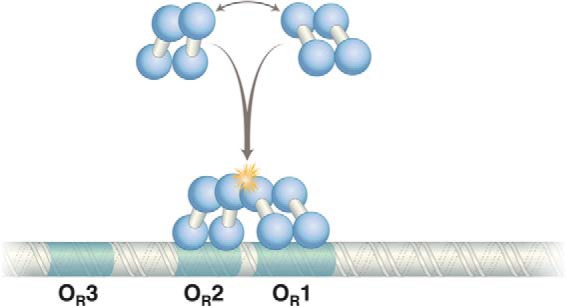
Cooperative binding of two λ repressor dimers to DNA. The N-terminal domains bind DNA, and contacts between C-terminal domains mediate dimerization and cooperative binding of dimers (starred) to DNA. The contact mediating the λ repressor N-terminal domain is separated from the C-terminal domain by a linker of 40 amino acids.
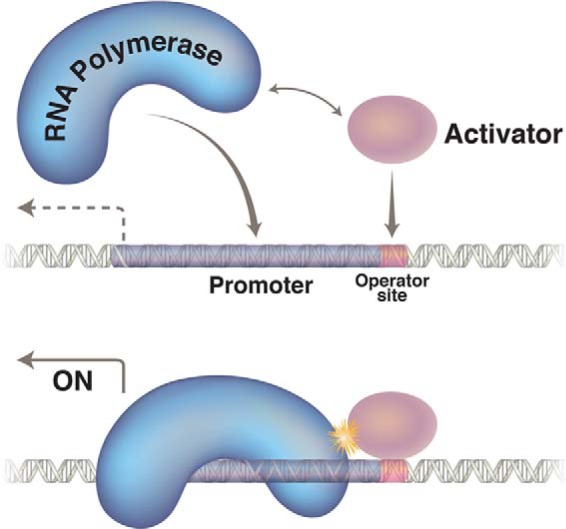
Transcription activation is a form of cooperative binding of proteins to DNA. The polymerase has a weak affinity for the promoter, and the activator (shown here binding to a single site) recruits it to a nearby promoter by virtue of a protein-protein contact.
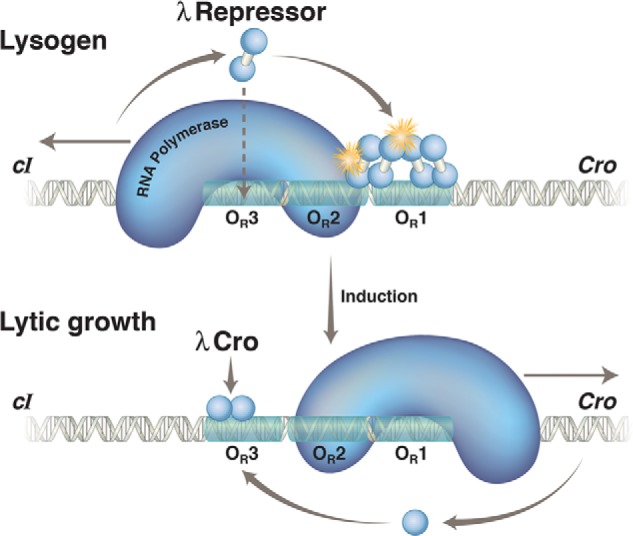
λ repressor and Cro action at OR in a lysogen and following induction.
Upper, in a lysogen, the λ repressor, encoded by the cI gene, preferentially occupies two adjacent sites, OR1 and OR2, in the operator OR. In this state, the repressor activates transcription of its own gene (which proceeds leftward in the figure) as it represses transcription of the cro gene (which would otherwise proceed rightward). With lower efficiency, the repressor also binds the weak site 3 and thereby turns off transcription of its own gene. Binding of the third site is facilitated by interaction with other repressor dimers bound to a site (called OL) some 3000 bp away, in an example of cooperative binding accommodated by DNA looping (not shown). The stars indicate protein-protein contacts of about equal strengths, one mediating cooperative binding of repressor dimers and the other recruitment of RNA polymerase and activation of transcription of the repressor gene. Lower, the repressor is destroyed (cleaved) upon exposure to UV irradiation. This relieves repression of cro and other genes required for lytic growth. Cro binds first to OR3 and abolishes transcription of the repressor gene. Later in the lytic cycle, Cro binds to sites 1 and 2 and represses transcription of its own gene (not shown). Not explicitly shown are the two relevant promoters, each of which covers ∼50 bp. PRM, which directs transcription of the cI gene in a lysogen, lies immediately adjacent to the operator site OR2, so the repressor and polymerase contact one another as they bind to their respective sites. In contrast, site OR3 overlaps this promoter, and hence, repressor bound there excludes polymerase. PR, which directs transcription of cro and other lytic genes, overlaps OR1 and OR2, so repressor bound to either site represses PR.
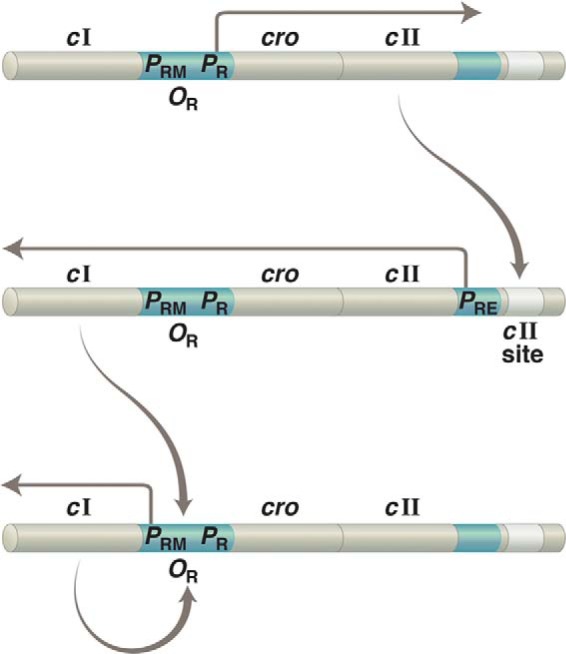
Establishment of lysogeny. The same gene, cI, is transcribed from two different promoters: from PRE to establish lysogeny and from PRM to maintain that state. Upper, upon infection of a bacterium with phage λ, transcription rightward from PR results in production of the transcriptional activator CII. Middle, CII binds to the “cII site” on DNA and activates leftward transcription from PRE. Lower, newly made repressor binds OR1 and OR2 and turns off transcription of cII (as well as of other genes required for lytic growth) as it simultaneously activates leftward transcription from PRE. As implied by this figure, PR controls not only lytic genes (as indicated in the text) but also cII, which is required to establish lysogeny. In addition, PR is an unusual promoter in that it requires no activator for full activity.
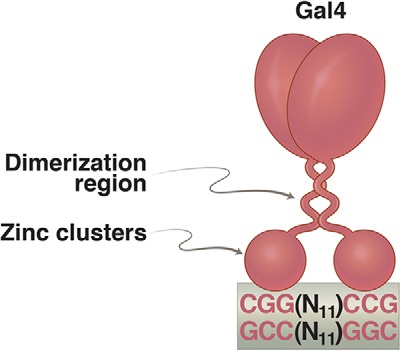
Gal4 dimer bound to a 17-bp 2-fold rotationally symmetric DNA site. Each of the “zinc clusters,” as they are called, recognizes a base pair triplet. These triplets are separated by 11 bp. Activators related to Gal4 bear identical zinc clusters, but the spacing between the triplets is unique in each case. The formal name for these zinc clusters is the Zn(II)Cys6 binuclear cluster. Such DNA-binding domains are not found in bacteria.
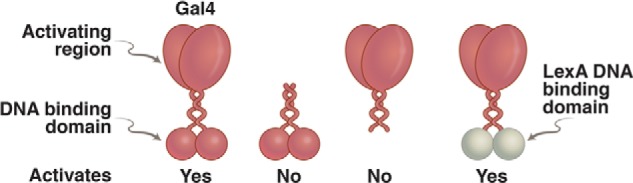
Domain-swap experiment. On the left is an intact Gal4, which binds DNA and activates transcription. To its right is the N-terminal domain alone, which dimerizes and binds DNA but cannot activate transcription. Second from the right is the separated C-terminal domain, which cannot bind DNA and cannot activate. On the far right is a hybrid protein in which the DNA-binding domain of the bacterial repressor LexA has been swapped for that of Gal4. This protein activates transcription in a wide array of eukaryotes, provided each target gene has been modified so as to bear a LexA-binding site nearby.
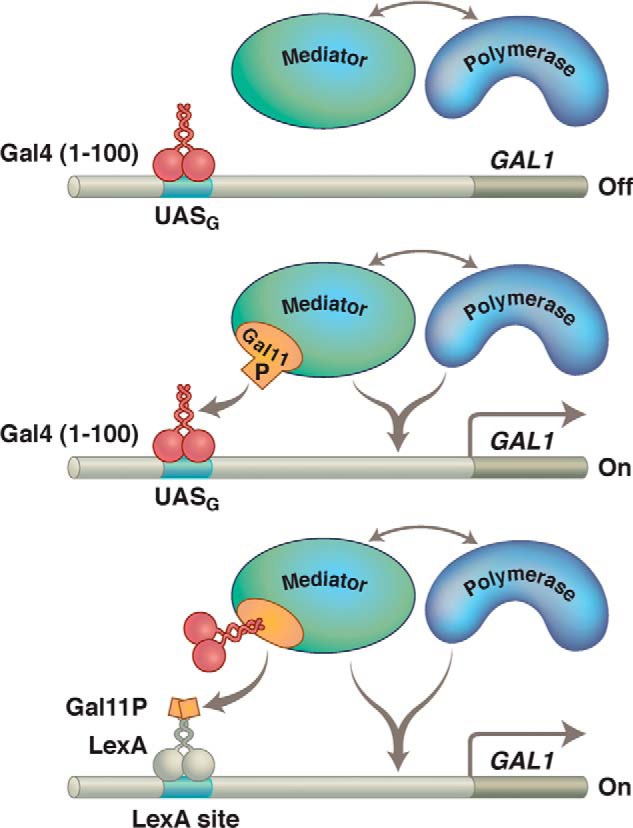
Gal11P effect. Gal11 is a component of the protein complex called the Mediator, shown here interacting with RNA polymerase. Upper, Gal4(1–100) (its N-terminal domain) does not contain an activating region and makes no contact with the polymerase-associated (i.e. Mediator) complex. This Gal11 fragment therefore does not activate transcription in a WT cell. Middle, the P (potentiator) mutation in the protein Gal11 creates a simple binding interaction with the Gal4 fragment, and that interaction (provided the Gal4 fragment is bound to DNA) results in activation of transcription. Lower, a bit of Gal11 bearing the P mutation has been fused to LexA, and that fragment activates only if the N-terminal domain of Gal4 has been fused to Gal11. There are many ways to effect recruitment.
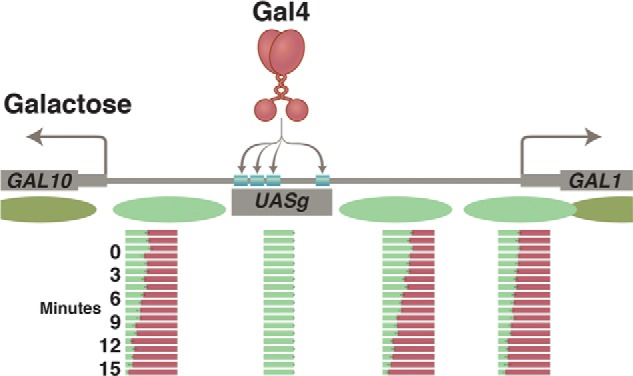
Nucleosomes at the GAL1/GAL10 genes before and after activation by Gal4. Prior to the addition of galactose, nucleosomes (green ovals) form in the promoters of the GAL1 and GAL10 genes. Upon addition of galactose, the exposed activating region of Gal4 recruits the enzyme SWI/SNF, which removes the promoter nucleosomes. The increasingly red bars show, at time points listed in minutes, the course of nucleosome removal. This step is followed by recruitment of the transcriptional machinery and transcription of both genes. Gal4 has acquired access to its four sites in the UASg, as explained in the text.
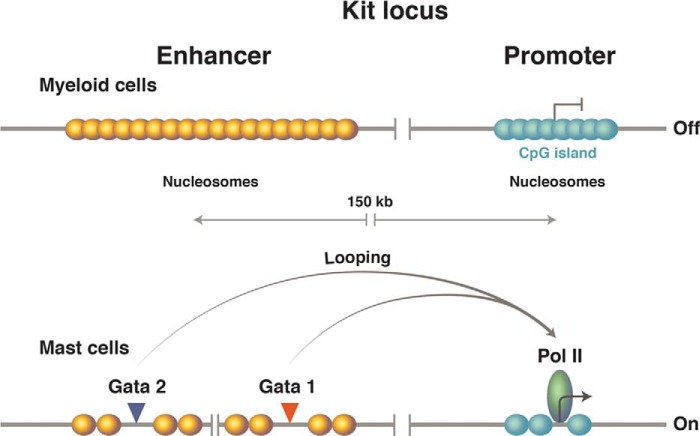
The murine kit locus.
Upper, the promoter, which lies in the CpG island, is covered by avidly forming nucleosomes. Formation of these promoter nucleosomes, we believe, ensures an extremely low basal level of transcription. The enhancer, positioned some 150 kb upstream, has a lower GC content, so the nucleosomes form there less avidly. Lower, specific DNA-binding proteins bind the enhancer, displacing enhancer nucleosomes. Then, accommodated by a giant DNA loop (not drawn), those DNA-bound activators replace one promoter nucleosome with the transcriptional machinery and thereby activate transcription. The remaining nucleosomes at the promoter now are phased passively thanks, we believe, to the barrier erected by the transcriptional machinery. (Only the presence of the polymerase is indicated at the promoter.) Pol II, polymerase II.
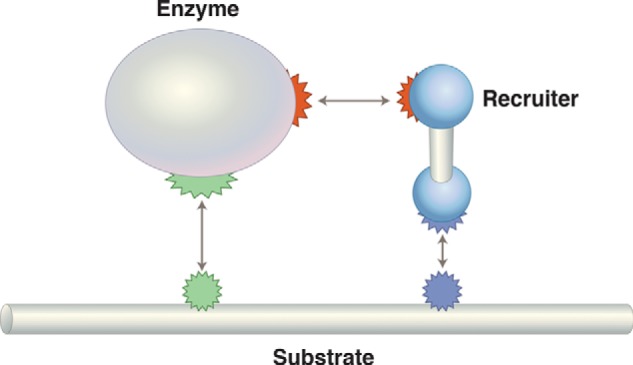
General regulatory scheme for regulation by recruitment. If the enzyme is RNA polymerase, then the recruiter is a transcriptional activator, and the substrate is DNA. To take another case, consider specific ubiquitylation: the enzyme would be an E2 ligase, the recruiter an E3 ligase, and the substrate a specific protein.
References
-
- Ptashne M. (2004) A Genetic Switch: Phage Lambda Revisited, 3rd Ed., Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY
-
- Ptashne M., Gann A. (2002) Genes & Signals, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY
-
- Ptashne M. (2013) François Jacob (1920–2013). Cell 153, 1180–1182 - PubMed
-
- Ptashne M. (1967) Specific binding of the lambda phage repressor to lambda DNA. Nature 214, 232–234 - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
