

Enhanced formation and disordered regulation of NETs in myeloperoxidase-ANCA-associated microscopic polyangiitis
- PMID: 24385592
- PMCID: PMC4005303
- DOI: 10.1681/ASN.2013060606
Enhanced formation and disordered regulation of NETs in myeloperoxidase-ANCA-associated microscopic polyangiitis
Abstract
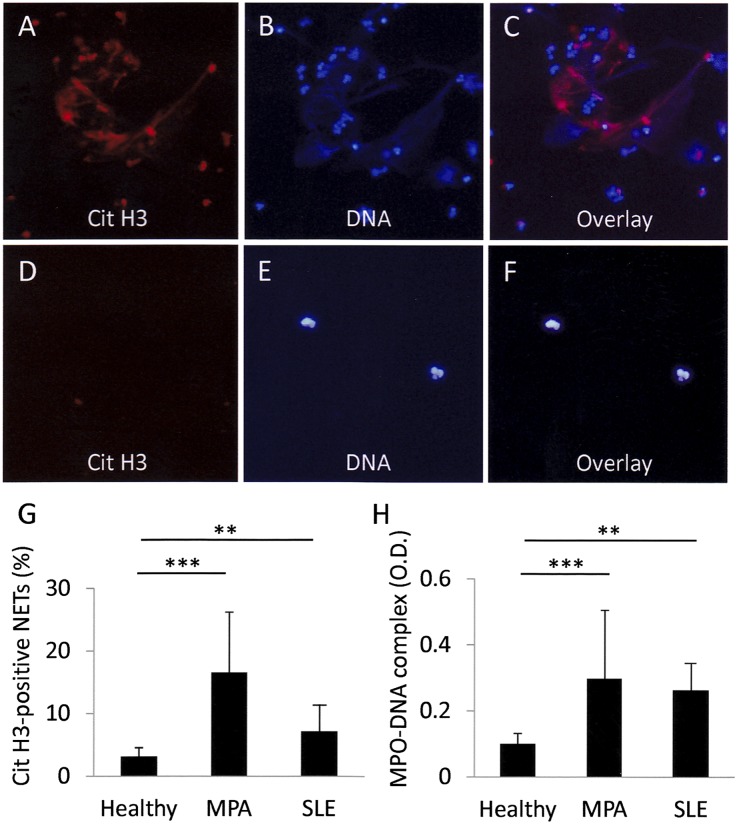
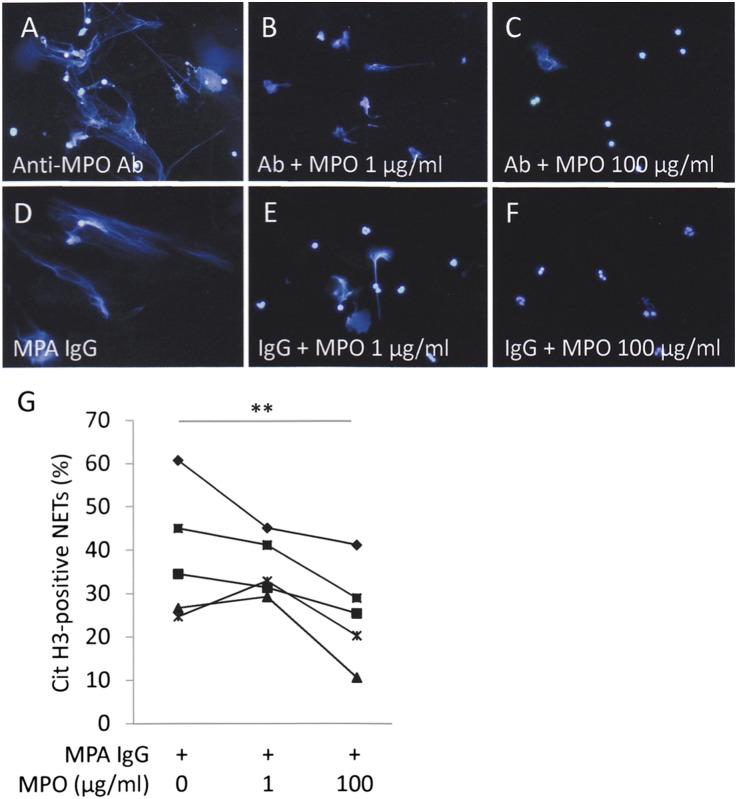
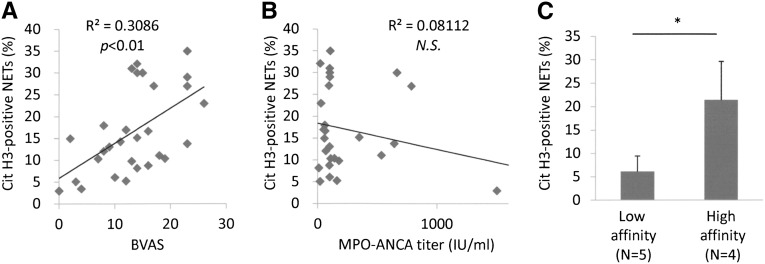
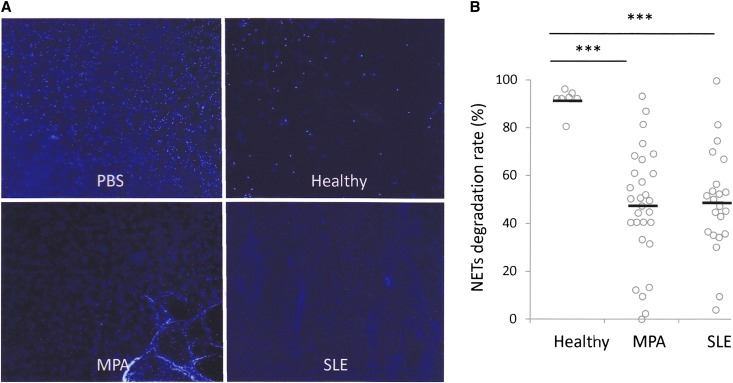
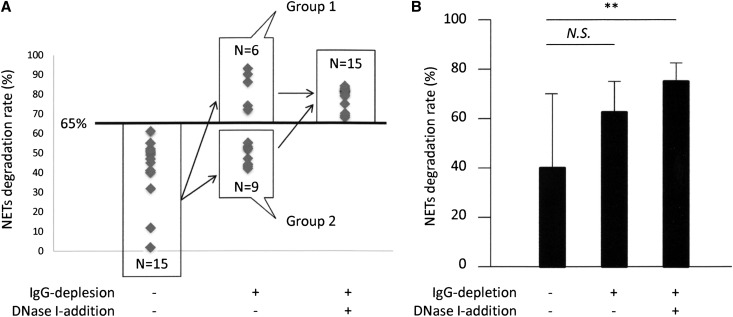
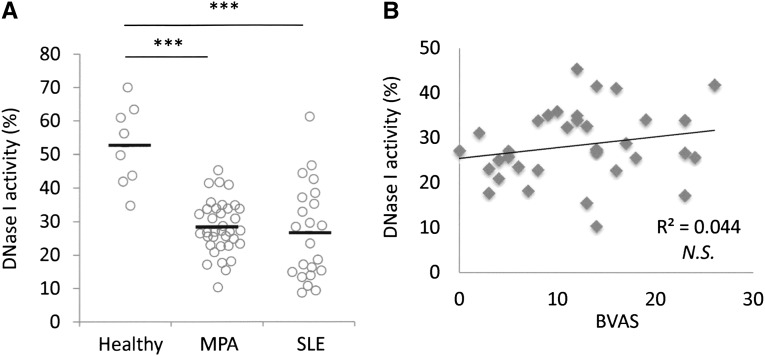
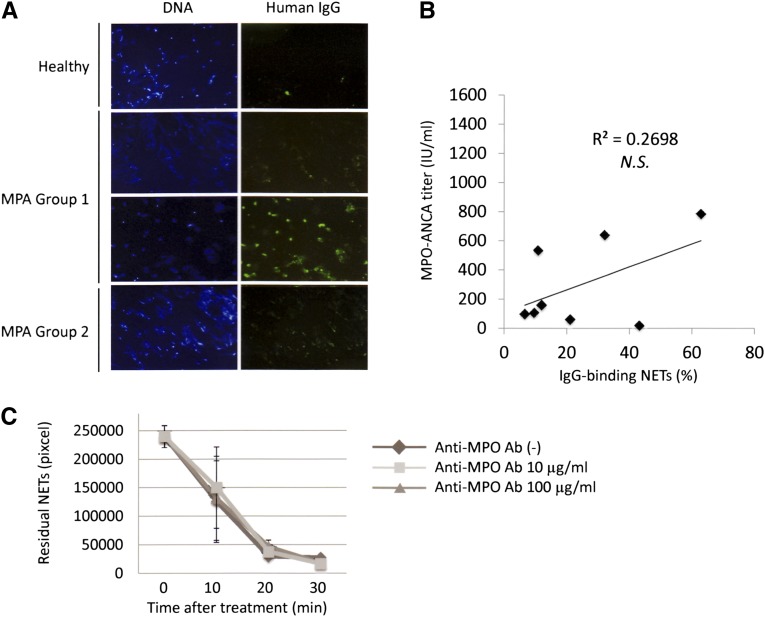
Microscopic polyangiitis (MPA) is an ANCA-associated vasculitis that affects small vessels, especially renal glomeruli. We recently demonstrated that the abnormal formation and impaired degradation of neutrophil extracellular traps (NETs) may be crucially involved in the generation of myeloperoxidase (MPO)-ANCA and subsequent development of MPA. This study assessed the formation and regulation of NETs in patients with MPO-ANCA-associated MPA. Peripheral blood samples were obtained from 38 patients with MPO-ANCA-associated MPA, 23 patients with systemic lupus erythematosus (SLE), and 8 healthy controls. IgG eluted from MPO-ANCA-associated MPA sera demonstrated the highest ability to induce NETs, and this ability correlated with disease activity and paralleled ANCA affinity for MPO. Moreover, addition of recombinant human MPO to these IgG samples reduced NET induction. Additionally, MPO-ANCA-associated MPA sera exhibited lower rates of NET degradation that recovered partially upon depletion of IgG. The activity of DNase I, an important regulator of NETs, was also lower in MPO-ANCA-associated MPA and SLE sera. IgG depletion from MPO-ANCA-associated MPA sera partially restored the rate of NET degradation, and addition of DNase I synergistically enhanced this restoration. Addition of anti-MPO antibodies did not inhibit DNase I activity, and some MPO-ANCA-associated MPA sera contained anti-NET antibodies at levels not correlated with MPO-ANCA titers, suggesting the involvement of unidentified autoantibodies as well. The collective evidence suggests a vicious cycle involving MPO-ANCA and the regulation of NETs could be critically involved in the pathogenesis of MPO-ANCA-associated MPA.
Keywords: ANCA; immunology; pathology; vasculitis.
Copyright © 2014 by the American Society of Nephrology.
Figures
References
-
- Jennette JC, Falk RJ, Bacon PA, Basu N, Cid MC, Ferrario F, Flores-Suarez LF, Gross WL, Guillevin L, Hagen EC, Hoffman GS, Jayne DR, Kallenberg CG, Lamprecht P, Langford CA, Luqmani RA, Mahr AD, Matteson EL, Merkel PA, Ozen S, Pusey CD, Rasmussen N, Rees AJ, Scott DG, Specks U, Stone JH, Takahashi K, Watts RA: 2012 revised International Chapel Hill Consensus Conference Nomenclature of Vasculitides. Arthritis Rheum 65: 1–11, 2013 - PubMed
-
- Csernok E: Anti-neutrophil cytoplasmic antibodies and pathogenesis of small vessel vasculitides. Autoimmun Rev 2: 158–164, 2003 - PubMed
-
- Chen M, Gao Y, Guo XH, Zhao MH: Propylthiouracil-induced antineutrophil cytoplasmic antibody-associated vasculitis. Nat Rev Nephrol 8: 476–483, 2012 - PubMed
-
- Nakazawa D, Tomaru U, Suzuki A, Masuda S, Hasegawa R, Kobayashi T, Nishio S, Kasahara M, Ishizu A: Abnormal conformation and impaired degradation of propylthiouracil-induced neutrophil extracellular traps: Implications of disordered neutrophil extracellular traps in a rat model of myeloperoxidase antineutrophil cytoplasmic antibody-associated vasculitis. Arthritis Rheum 64: 3779–3787, 2012 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
