

Cyclin A degradation by primate cytomegalovirus protein pUL21a counters its innate restriction of virus replication
- PMID: 24385906
- PMCID: PMC3873445
- DOI: 10.1371/journal.ppat.1003825
Cyclin A degradation by primate cytomegalovirus protein pUL21a counters its innate restriction of virus replication
Abstract
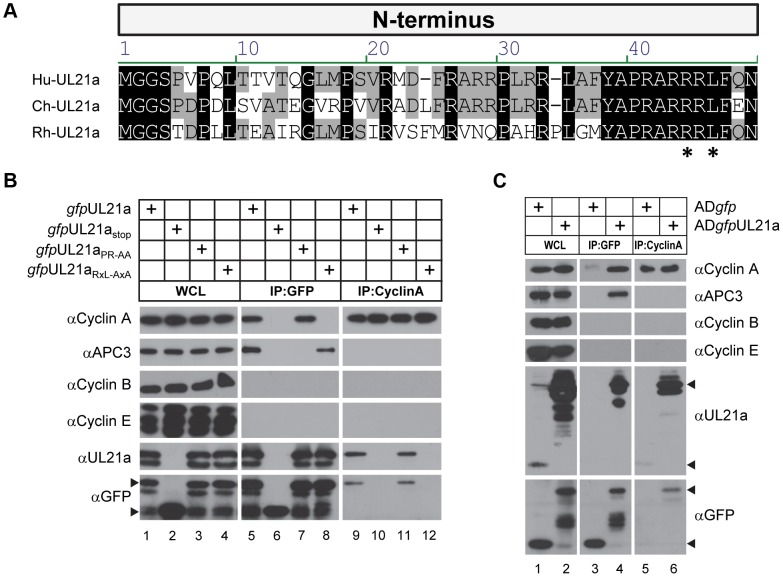
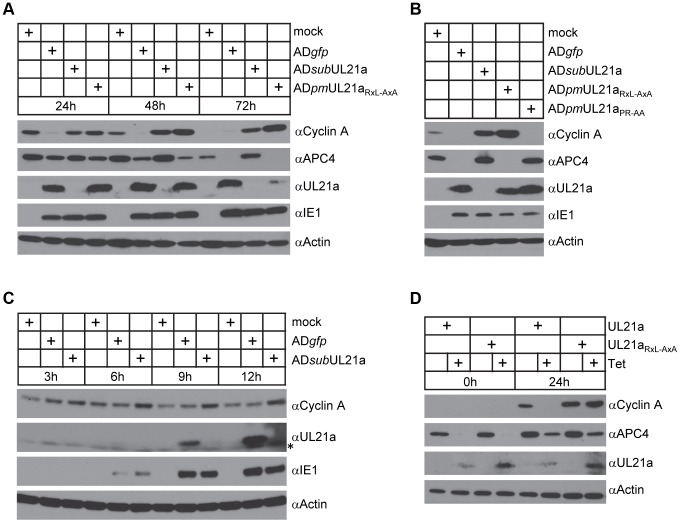
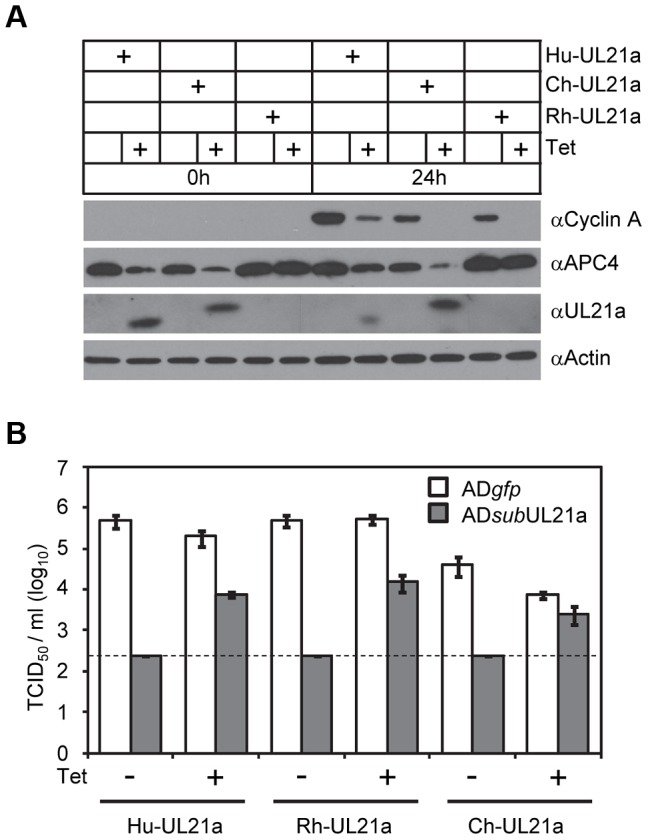
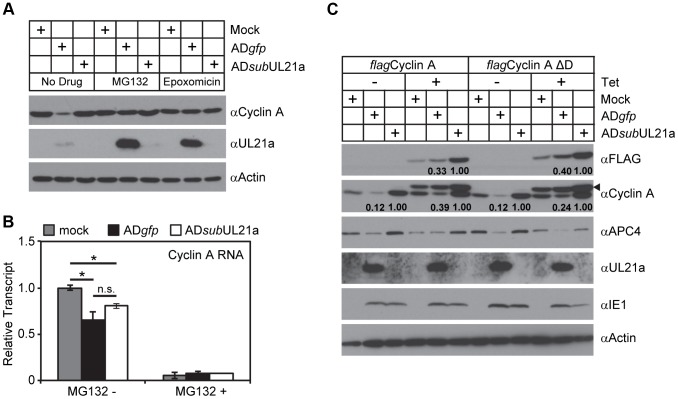
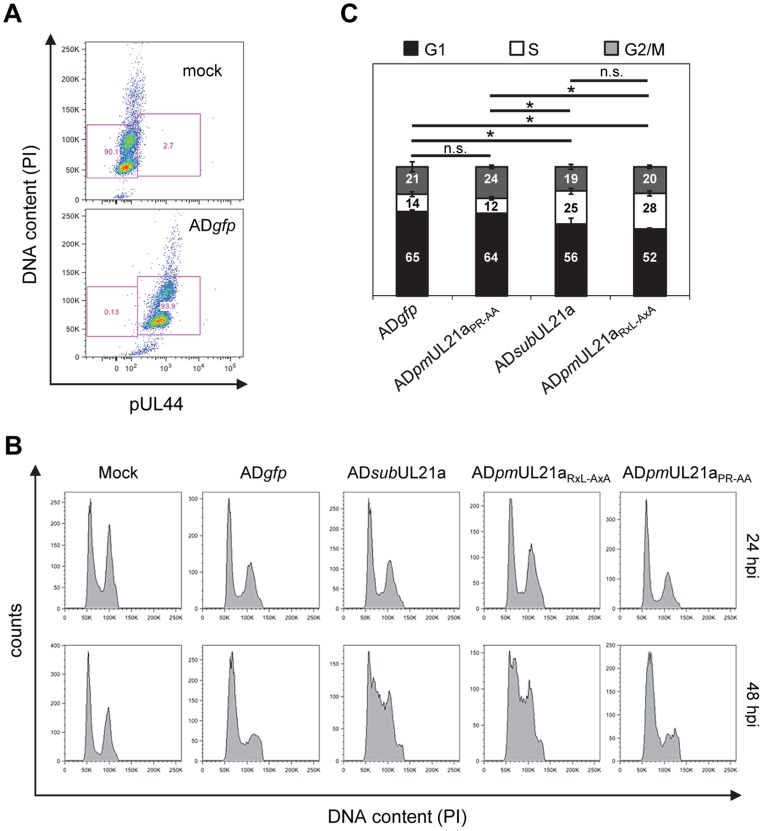
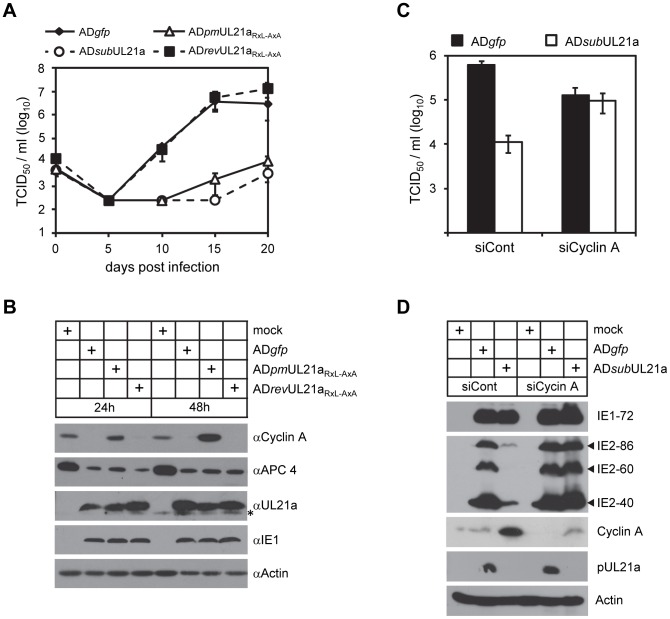
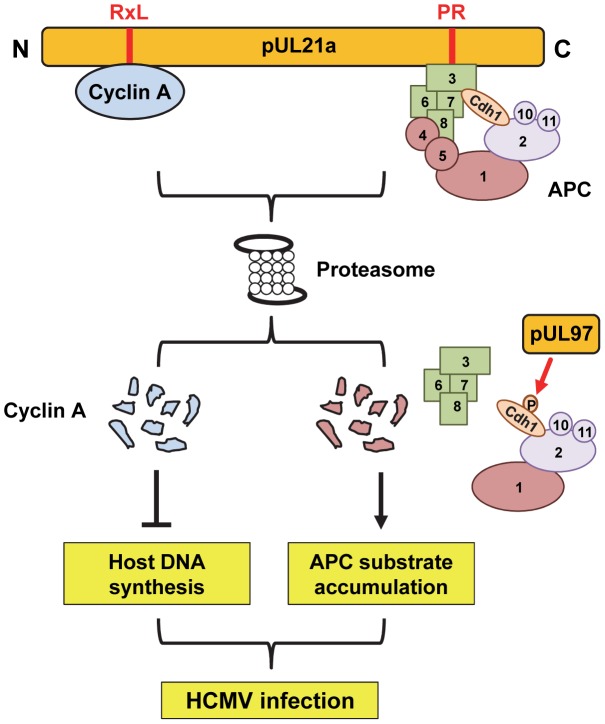
Cyclin A is critical for cellular DNA synthesis and S phase progression of the cell cycle. Human cytomegalovirus (HCMV) can reduce cyclin A levels and block cellular DNA synthesis, and cyclin A overexpression can repress HCMV replication. This interaction has only been previously observed in HCMV as murine CMV does not downregulate cyclin A, and the responsible viral factor has not been identified. We previously reported that the HCMV protein pUL21a disrupted the anaphase-promoting complex (APC), but a point mutant abrogating this activity did not phenocopy a UL21a-deficient virus, suggesting that pUL21a has an additional function. Here we identified a conserved arginine-x-leucine (RxL) cyclin-binding domain within pUL21a, which allowed pUL21a to interact with cyclin A and target it for proteasome degradation. Homologous pUL21a proteins from both chimpanzee and rhesus CMVs also contained the RxL domain and similarly degraded cyclin A, indicating that this function is conserved in primate CMVs. The RxL point mutation disabled the virus' ability to block cellular DNA synthesis and resulted in a growth defect similar to pUL21a-deficient virus. Importantly, knockdown of cyclin A rescued growth of UL21a-deficient virus. Together, these data show that during evolution, the pUL21a family proteins of primate CMVs have acquired a cyclin-binding domain that targets cyclin A for degradation, thus neutralizing its restriction on virus replication. Finally, the combined proteasome-dependent degradation of pUL21a and its cellular targets suggests that pUL21a may act as a novel suicide protein, targeting its protein cargos for destruction.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
