

Transcriptome analysis in cotton boll weevil (Anthonomus grandis) and RNA interference in insect pests
- PMID: 24386449
- PMCID: PMC3874031
- DOI: 10.1371/journal.pone.0085079
Transcriptome analysis in cotton boll weevil (Anthonomus grandis) and RNA interference in insect pests
Abstract
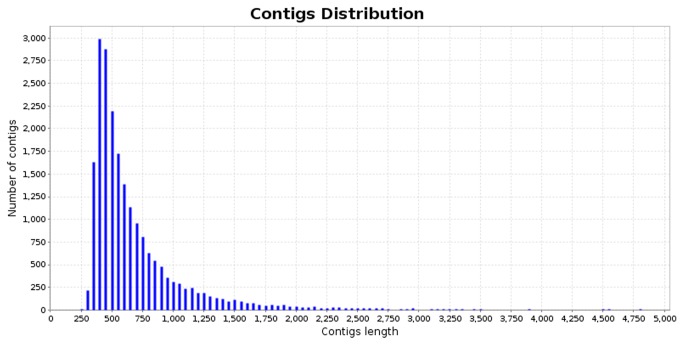
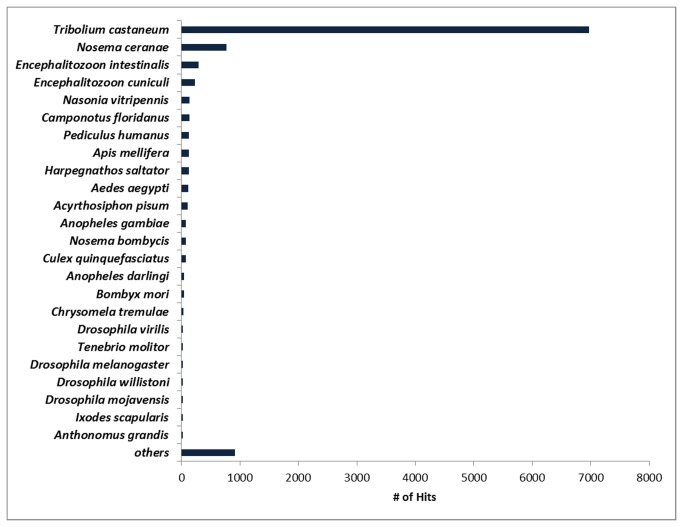
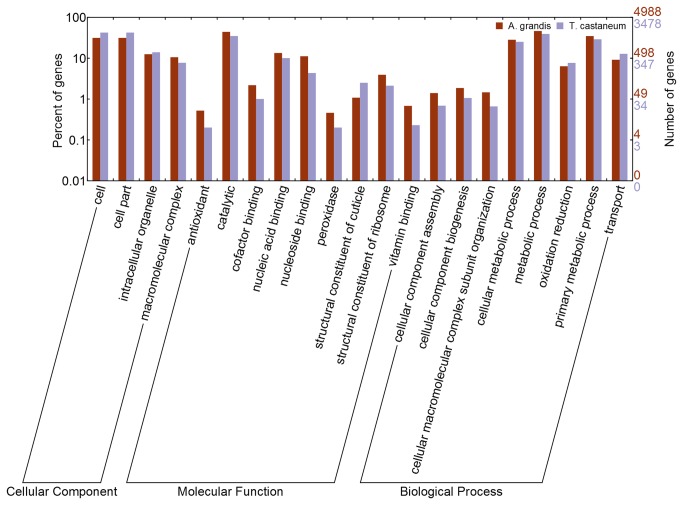
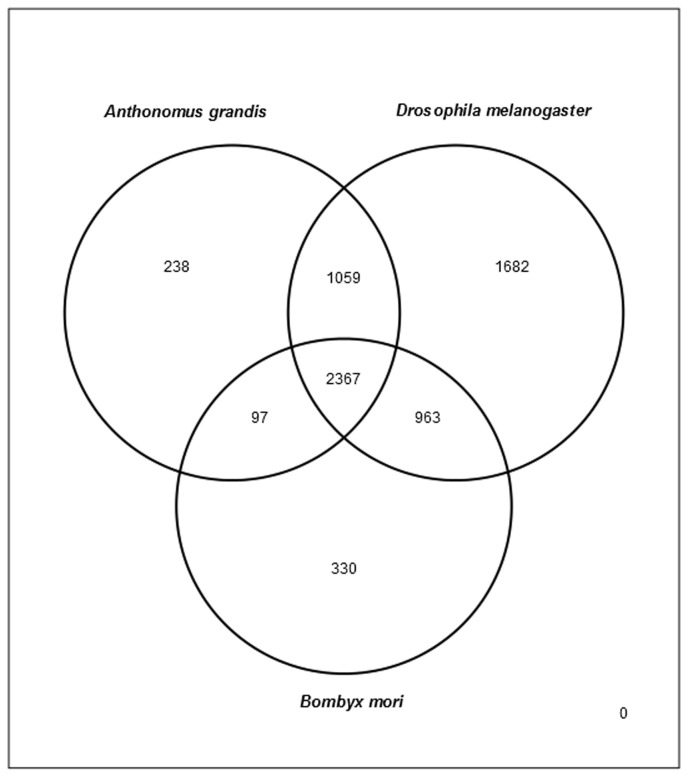
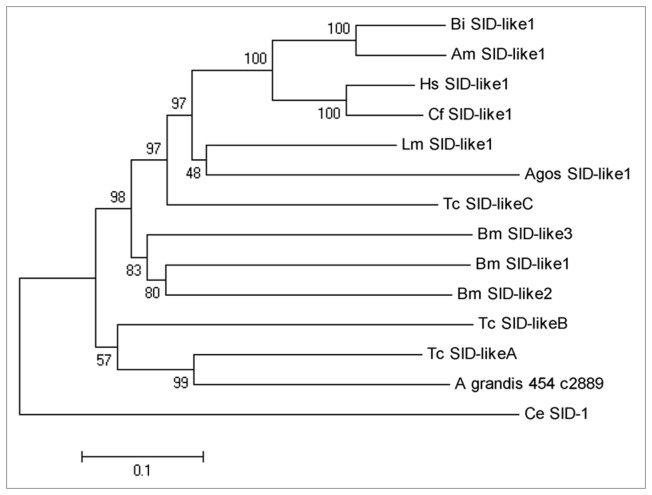
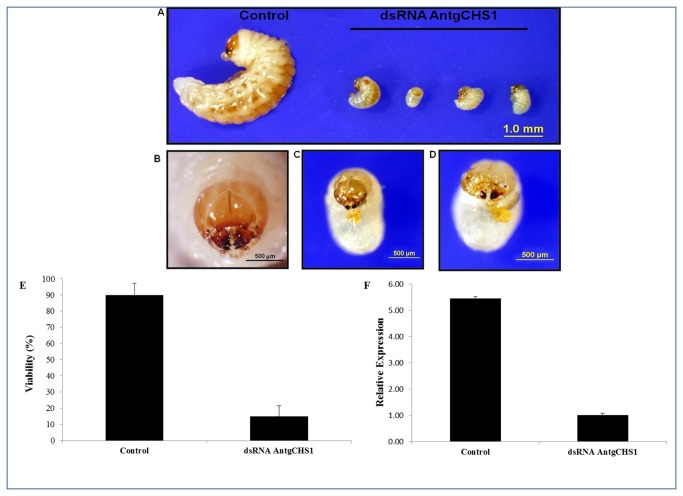
Cotton plants are subjected to the attack of several insect pests. In Brazil, the cotton boll weevil, Anthonomus grandis, is the most important cotton pest. The use of insecticidal proteins and gene silencing by interference RNA (RNAi) as techniques for insect control are promising strategies, which has been applied in the last few years. For this insect, there are not much available molecular information on databases. Using 454-pyrosequencing methodology, the transcriptome of all developmental stages of the insect pest, A. grandis, was analyzed. The A. grandis transcriptome analysis resulted in more than 500.000 reads and a data set of high quality 20,841 contigs. After sequence assembly and annotation, around 10,600 contigs had at least one BLAST hit against NCBI non-redundant protein database and 65.7% was similar to Tribolium castaneum sequences. A comparison of A. grandis, Drosophila melanogaster and Bombyx mori protein families' data showed higher similarity to dipteran than to lepidopteran sequences. Several contigs of genes encoding proteins involved in RNAi mechanism were found. PAZ Domains sequences extracted from the transcriptome showed high similarity and conservation for the most important functional and structural motifs when compared to PAZ Domains from 5 species. Two SID-like contigs were phylogenetically analyzed and grouped with T. castaneum SID-like proteins. No RdRP gene was found. A contig matching chitin synthase 1 was mined from the transcriptome. dsRNA microinjection of a chitin synthase gene to A. grandis female adults resulted in normal oviposition of unviable eggs and malformed alive larvae that were unable to develop in artificial diet. This is the first study that characterizes the transcriptome of the coleopteran, A. grandis. A new and representative transcriptome database for this insect pest is now available. All data support the state of the art of RNAi mechanism in insects.
Conflict of interest statement
Figures


References
-
- Consortium ikI (2012) 5,000 Insect Genome Project. Arthropod Genomes
-
- Legrand S, Valot N, Nicolé F, Moja S, Baudino S et al. (2010) One-step identification of conserved miRNAs, their targets, potential transcription factors and effector genes of complete secondary metabolism pathways after 454 pyrosequencing of calyx cDNAs from the Labiate Salvia sclarea L. Gene 450: 55-62. doi: 10.1016/j.gene.2009.10.004. PubMed: 19840835. - DOI - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
