

Multifarious determinants of cytokine receptor signaling specificity
- PMID: 24388212
- PMCID: PMC4449261
- DOI: 10.1016/B978-0-12-800100-4.00001-5
Multifarious determinants of cytokine receptor signaling specificity
Abstract
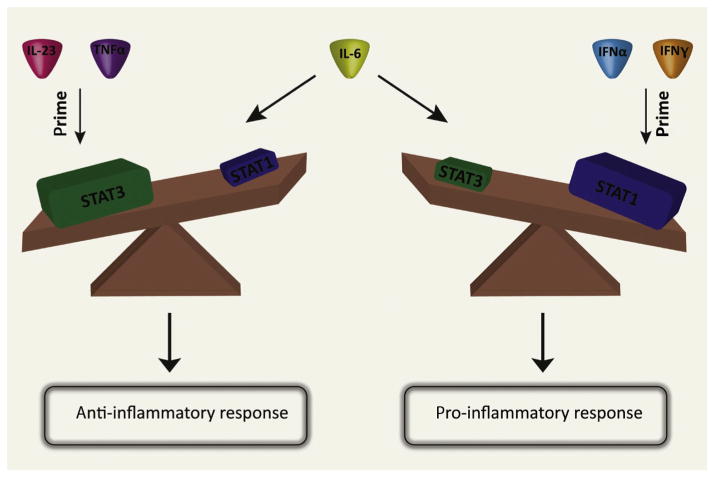
Cytokines play crucial roles in regulating immune homeostasis. Two important characteristics of most cytokines are pleiotropy, defined as the ability of one cytokine to exhibit diverse functionalities, and redundancy, defined as the ability of multiple cytokines to exert overlapping activities. Identifying the determinants for unique cellular responses to cytokines in the face of shared receptor usage, pleiotropy, and redundancy will be essential in order to harness the potential of cytokines as therapeutics. Here, we discuss the biophysical (ligand-receptor geometry and affinity) and cellular (receptor trafficking and intracellular abundance of signaling molecules) parameters that contribute to the specificity of cytokine bioactivities. Whereas the role of extracellular ternary complex geometry in cytokine-induced signaling is still not completely elucidated, cytokine-receptor affinity is known to impact signaling through modulation of the stability and kinetics of ternary complex formation. Receptor trafficking also plays an important and likely underappreciated role in the diversification of cytokine bioactivities but it has been challenging to experimentally probe trafficking effects. We also review recent efforts to quantify levels of intracellular signaling components, as second messenger abundance can affect cytokine-induced bioactivities both quantitatively and qualitatively. We conclude by discussing the application of protein engineering to develop therapeutically relevant cytokines with reduced pleiotropy and redirected biological functionalities.
Keywords: Cytokine engineering; Cytokine signaling; Cytokine structure; Functional specificity; Interleukin; Ligand–receptor affinity; Receptor trafficking.
© 2014 Elsevier Inc. All rights reserved.
Figures


Similar articles
-
Mapping Determinants of Cytokine Signaling via Protein Engineering.Front Immunol. 2018 Sep 27;9:2143. doi: 10.3389/fimmu.2018.02143. eCollection 2018. Front Immunol. 2018. PMID: 30319612 Free PMC article. Review.
-
Instructive roles for cytokine-receptor binding parameters in determining signaling and functional potency.Sci Signal. 2015 Nov 10;8(402):ra114. doi: 10.1126/scisignal.aab2677. Sci Signal. 2015. PMID: 26554818 Free PMC article.
-
Cytokine pleiotropy and redundancy: a view from the receptor.Stem Cells. 1994;12 Suppl 1:3-12; discussion 12-4. Stem Cells. 1994. PMID: 7696967 Review.
-
Insights into cytokine-receptor interactions from cytokine engineering.Annu Rev Immunol. 2015;33:139-67. doi: 10.1146/annurev-immunol-032713-120211. Epub 2014 Dec 10. Annu Rev Immunol. 2015. PMID: 25493332 Free PMC article. Review.
-
Two different epitopes of the signal transducer gp130 sequentially cooperate on IL-6-induced receptor activation.J Immunol. 2000 Dec 15;165(12):7042-9. doi: 10.4049/jimmunol.165.12.7042. J Immunol. 2000. PMID: 11120832
Cited by
-
Engineering a Single-Agent Cytokine/Antibody Fusion That Selectively Expands Regulatory T Cells for Autoimmune Disease Therapy.J Immunol. 2018 Oct 1;201(7):2094-2106. doi: 10.4049/jimmunol.1800578. Epub 2018 Aug 13. J Immunol. 2018. PMID: 30104245 Free PMC article.
-
Four-color single-molecule imaging with engineered tags resolves the molecular architecture of signaling complexes in the plasma membrane.Cell Rep Methods. 2022 Feb 4;2(2):100165. doi: 10.1016/j.crmeth.2022.100165. eCollection 2022 Feb 28. Cell Rep Methods. 2022. PMID: 35474965 Free PMC article.
-
Mapping Determinants of Cytokine Signaling via Protein Engineering.Front Immunol. 2018 Sep 27;9:2143. doi: 10.3389/fimmu.2018.02143. eCollection 2018. Front Immunol. 2018. PMID: 30319612 Free PMC article. Review.
-
Possibility of averting cytokine storm in SARS-COV 2 patients using specialized pro-resolving lipid mediators.Biochem Pharmacol. 2023 Mar;209:115437. doi: 10.1016/j.bcp.2023.115437. Epub 2023 Jan 30. Biochem Pharmacol. 2023. PMID: 36731803 Free PMC article. Review.
-
Topological control of cytokine receptor signaling induces differential effects in hematopoiesis.Science. 2019 May 24;364(6442):eaav7532. doi: 10.1126/science.aav7532. Epub 2019 May 23. Science. 2019. PMID: 31123111 Free PMC article.
References
-
- Andrews AL, Holloway JW, Holgate ST, Davies DE. IL-4 receptor alpha is an important modulator of IL-4 and IL-13 receptor binding: Implications for the development of therapeutic targets. Journal of Immunology. 2006;176:7456–7461. - PubMed
-
- Andrews AL, Holloway JW, Puddicombe SM, Holgate ST, Davies DE. Kinetic analysis of the interleukin-13 receptor complex. Journal of Biological Chemistry. 2002;277:46073–46078. - PubMed
-
- Avalos BR, Gasson JC, Hedvat C, Quan SG, Baldwin GC, Weisbart RH, et al. Human granulocyte colony-stimulating factor: Biologic activities and receptor characterization on hematopoietic cells and small cell lung cancer cell lines. Blood. 1990;75:851–857. - PubMed
-
- Bagley CJ, Woodcock JM, Hercus TR, Shannon MF, Lopez AF. Interaction of GM-CSF and IL-3 with the common beta-chain of their receptors. Journal of Leukocyte Biology. 1995;57:739–746. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
