

Structure-based mechanism for Na(+)/melibiose symport by MelB
- PMID: 24389923
- PMCID: PMC4026327
- DOI: 10.1038/ncomms4009
Structure-based mechanism for Na(+)/melibiose symport by MelB
Abstract
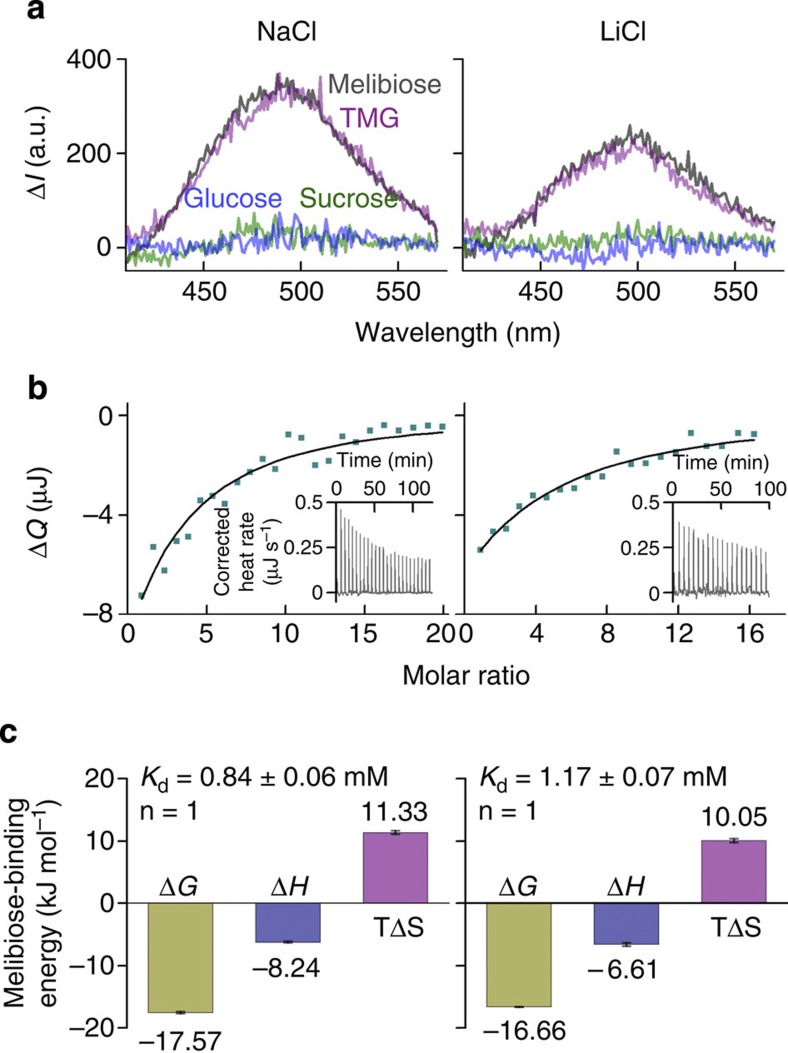
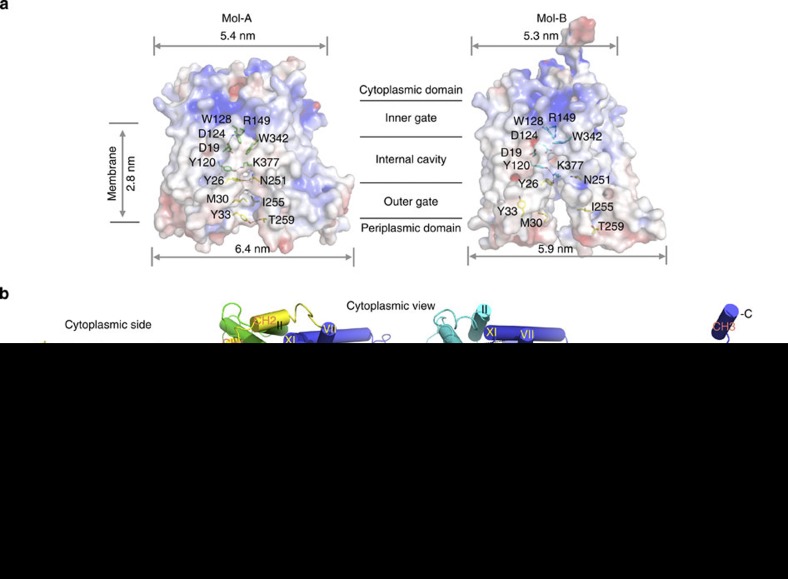
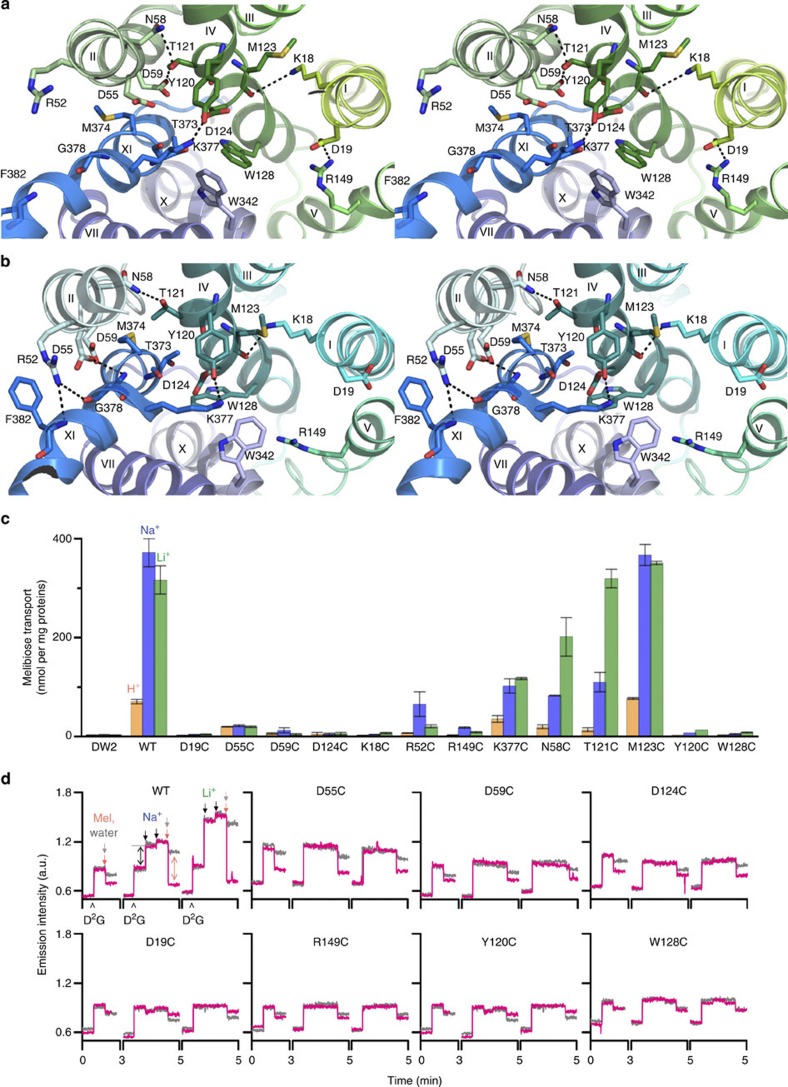
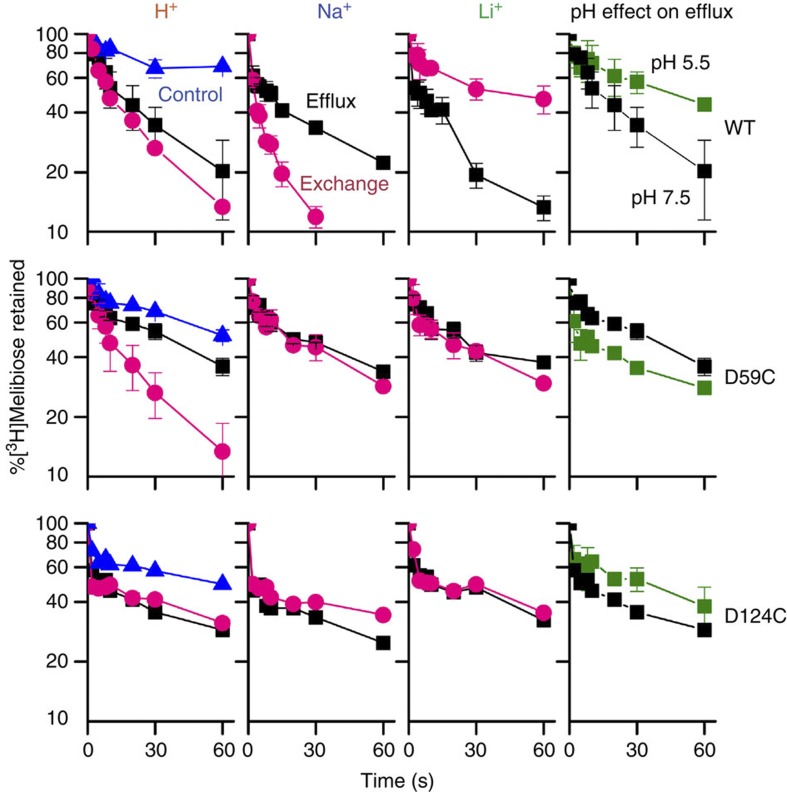
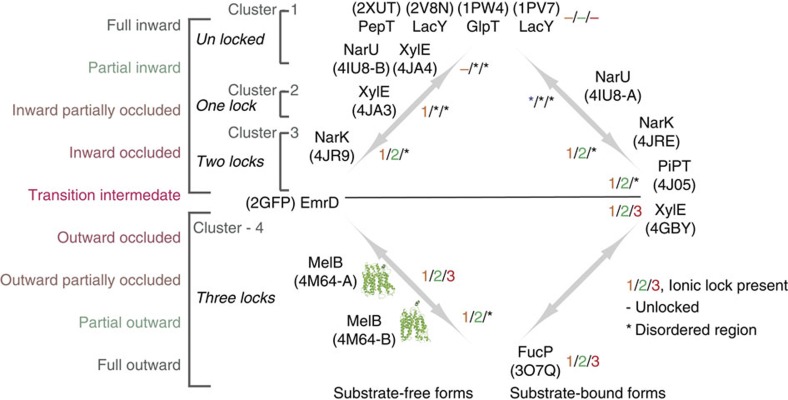
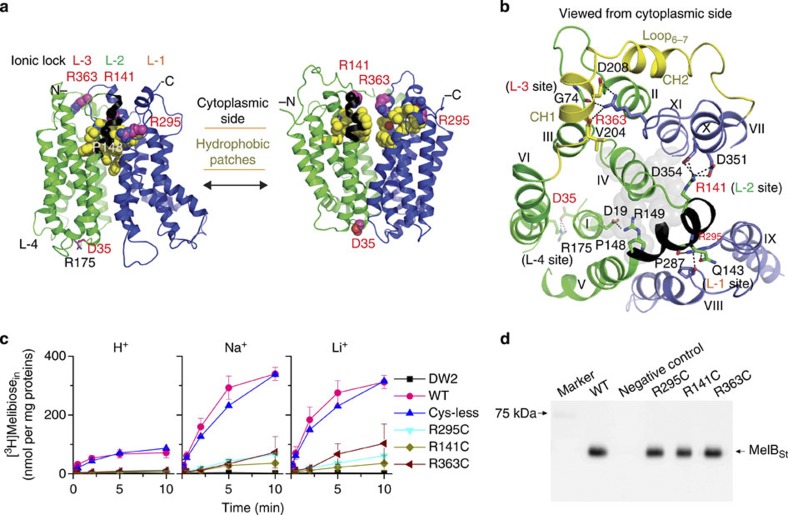
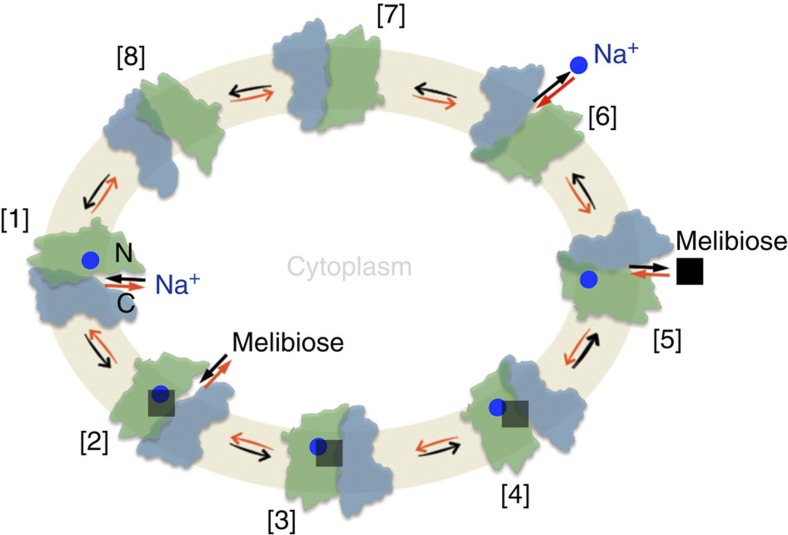
The bacterial melibiose permease (MelB) belongs to the glycoside-pentoside-hexuronide:cation symporter family, a part of the major facilitator superfamily (MFS). Structural information regarding glycoside-pentoside-hexuronide:cation symporter family transporters and other Na(+)-coupled permeases within MFS has been lacking, although a wealth of biochemical and biophysical data are available. Here we present the three-dimensional crystal structures of Salmonella typhimurium MelBSt in two conformations, representing an outward partially occluded and an outward inactive state of MelBSt. MelB adopts a typical MFS fold and contains a previously unidentified cation-binding motif. Three conserved acidic residues form a pyramidal-shaped cation-binding site for Na(+), Li(+) or H(+), which is in close proximity to the sugar-binding site. Both cosubstrate-binding sites are mainly contributed by the residues from the amino-terminal domain. These two structures and the functional data presented here provide mechanistic insights into Na(+)/melibiose symport. We also postulate a structural foundation for the conformational cycling necessary for transport catalysed by MFS permeases in general.
Figures
References
-
- Poolman B. et al.. Cation and sugar selectivity determinants in a novel family of transport proteins. Mol. Microbiol. 19, 911–922 (1996). - PubMed
-
- Saier M. H. Jr. Families of transmembrane sugar transport proteins. Mol. Microbiol. 35, 699–710 (2000). - PubMed
-
- Mizushima K. et al.. Cloning and sequencing of the melB gene encoding the melibiose permease of Salmonella typhimurium LT2. Mol. Gen. Genet. 234, 74–80 (1992). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
