

Flies and humans share a motion estimation strategy that exploits natural scene statistics
- PMID: 24390225
- PMCID: PMC3993001
- DOI: 10.1038/nn.3600
Flies and humans share a motion estimation strategy that exploits natural scene statistics
Abstract
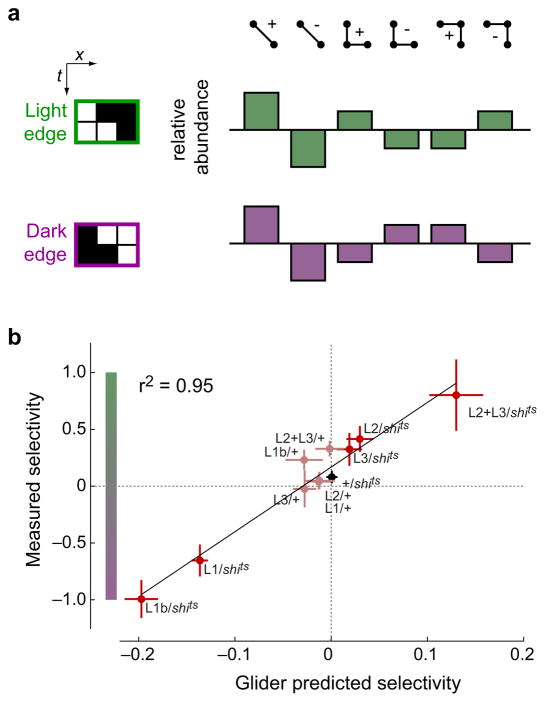
Sighted animals extract motion information from visual scenes by processing spatiotemporal patterns of light falling on the retina. The dominant models for motion estimation exploit intensity correlations only between pairs of points in space and time. Moving natural scenes, however, contain more complex correlations. We found that fly and human visual systems encode the combined direction and contrast polarity of moving edges using triple correlations that enhance motion estimation in natural environments. Both species extracted triple correlations with neural substrates tuned for light or dark edges, and sensitivity to specific triple correlations was retained even as light and dark edge motion signals were combined. Thus, both species separately process light and dark image contrasts to capture motion signatures that can improve estimation accuracy. This convergence argues that statistical structures in natural scenes have greatly affected visual processing, driving a common computational strategy over 500 million years of evolution.
Figures






References
-
- Field DJ. Relations between the statistics of natural images and the response properties of cortical cells. J Opt Soc Am A. 1987;4:2379–2394. - PubMed
-
- Ruderman DL, Bialek W. Statistics of natural images: Scaling in the woods. Physical Review Letters. 1994;73:814–817. - PubMed
-
- Simoncelli EP, Olshausen BA. Natural image statistics and neural representation. Annual Review of Neuroscience. 2001;24:1193–1216. - PubMed
-
- Hassenstein B, Reichardt W. Systemtheoretische Analyse der Zeit-, Reihenfolgen-und Vorzeichenauswertung bei der Bewegungsperzeption des Rüsselkäfers Chlorophanus. Zeitschrift für Naturforschung. 1956;11:513–524.
-
- Adelson E, Bergen J. Spatiotemporal energy models for the perception of motion. Journal of the Optical Society of America A. 1985;2:284–299. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
