

The velvet family of fungal regulators contains a DNA-binding domain structurally similar to NF-κB
- PMID: 24391470
- PMCID: PMC3876986
- DOI: 10.1371/journal.pbio.1001750
The velvet family of fungal regulators contains a DNA-binding domain structurally similar to NF-κB
Erratum in
- PLoS Biol. 2014 Apr;12(4):e1001849
Abstract
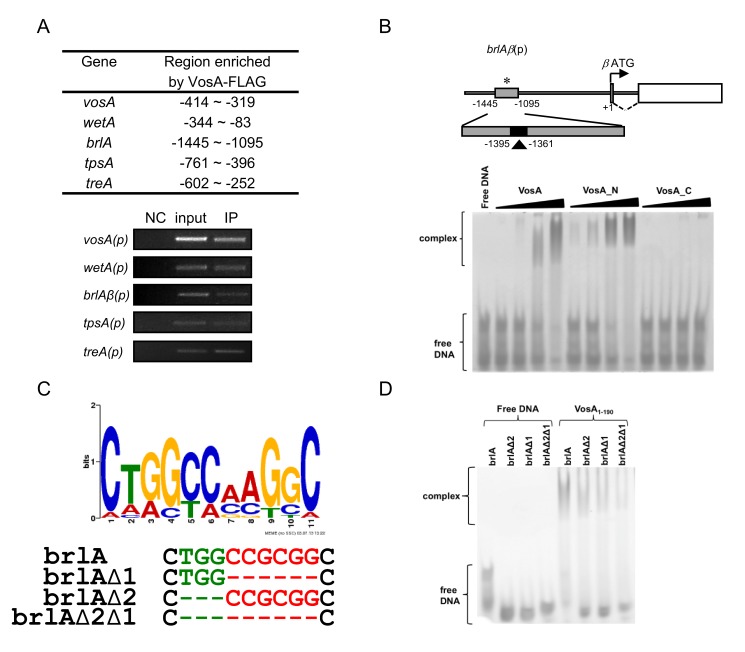
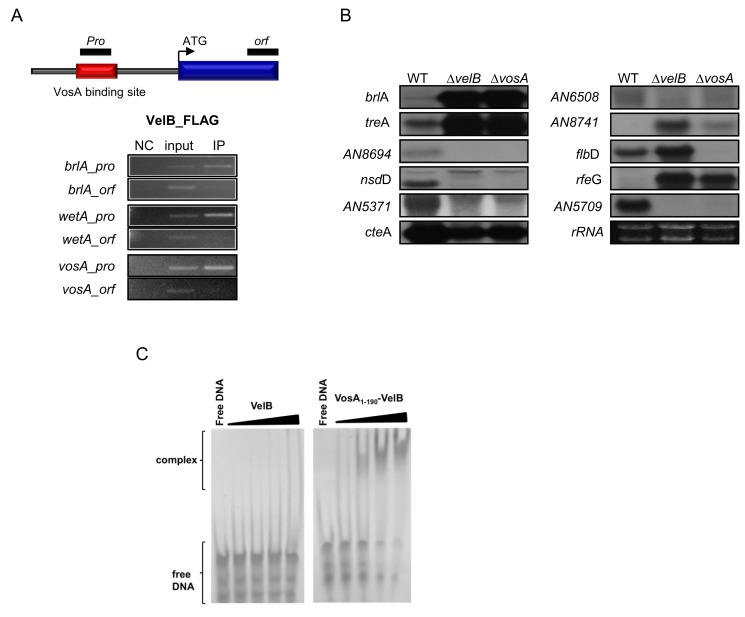
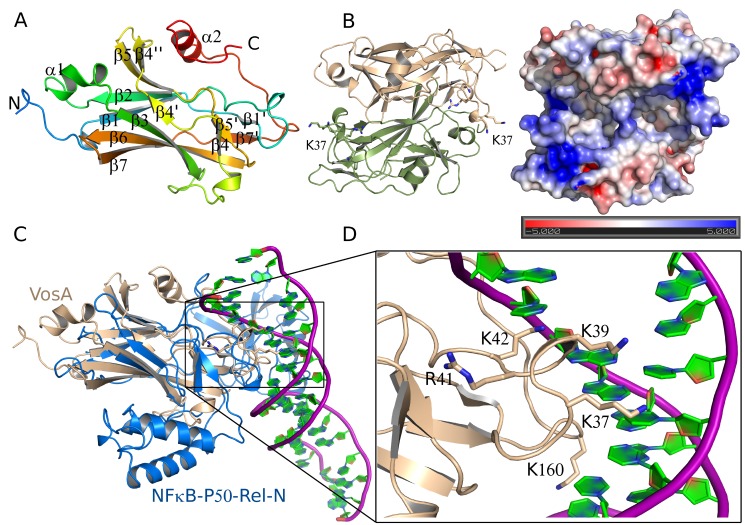
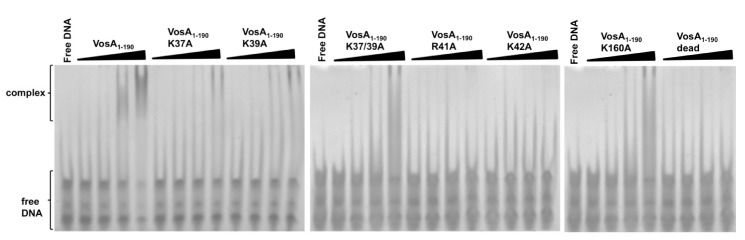
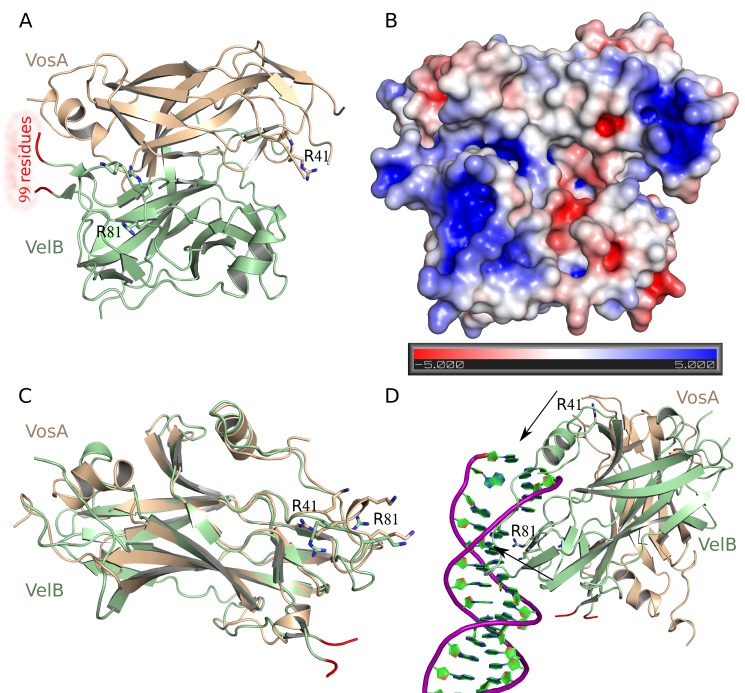
Morphological development of fungi and their combined production of secondary metabolites are both acting in defence and protection. These processes are mainly coordinated by velvet regulators, which contain a yet functionally and structurally uncharacterized velvet domain. Here we demonstrate that the velvet domain of VosA is a novel DNA-binding motif that specifically recognizes an 11-nucleotide consensus sequence consisting of two motifs in the promoters of key developmental regulatory genes. The crystal structure analysis of the VosA velvet domain revealed an unforeseen structural similarity with the Rel homology domain (RHD) of the mammalian transcription factor NF-κB. Based on this structural similarity several conserved amino acid residues present in all velvet domains have been identified and shown to be essential for the DNA binding ability of VosA. The velvet domain is also involved in dimer formation as seen in the solved crystal structures of the VosA homodimer and the VosA-VelB heterodimer. These findings suggest that defence mechanisms of both fungi and animals might be governed by structurally related DNA-binding transcription factors.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
Comment in
-
The velvet underground emerges.PLoS Biol. 2013 Dec;11(12):e1001751. doi: 10.1371/journal.pbio.1001751. Epub 2013 Dec 31. PLoS Biol. 2013. PMID: 24391471 Free PMC article. No abstract available.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
