

Inflammation fuels colicin Ib-dependent competition of Salmonella serovar Typhimurium and E. coli in enterobacterial blooms
- PMID: 24391500
- PMCID: PMC3879352
- DOI: 10.1371/journal.ppat.1003844
Inflammation fuels colicin Ib-dependent competition of Salmonella serovar Typhimurium and E. coli in enterobacterial blooms
Abstract
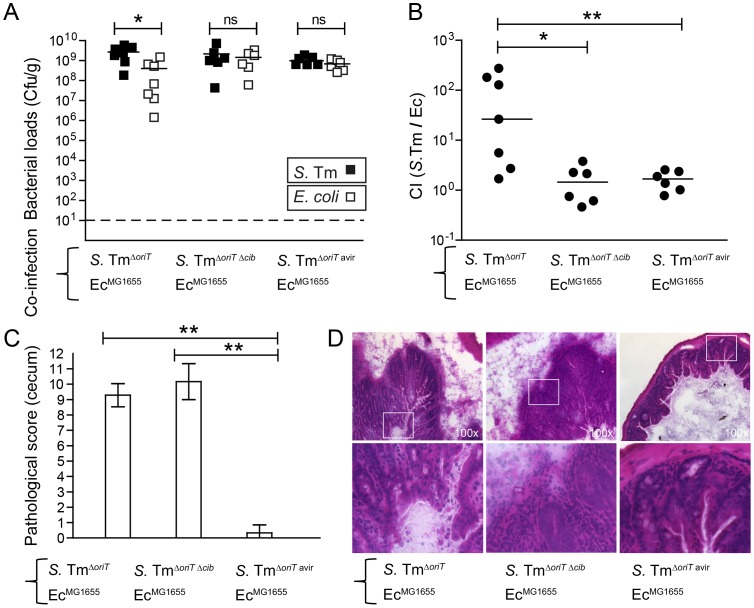
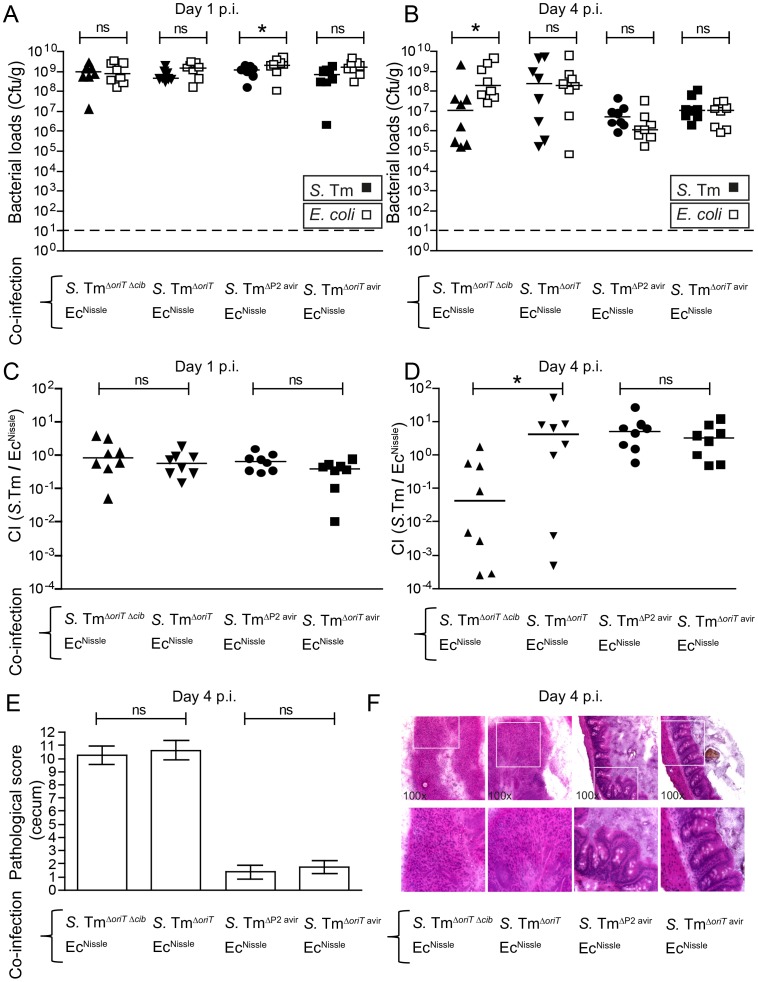
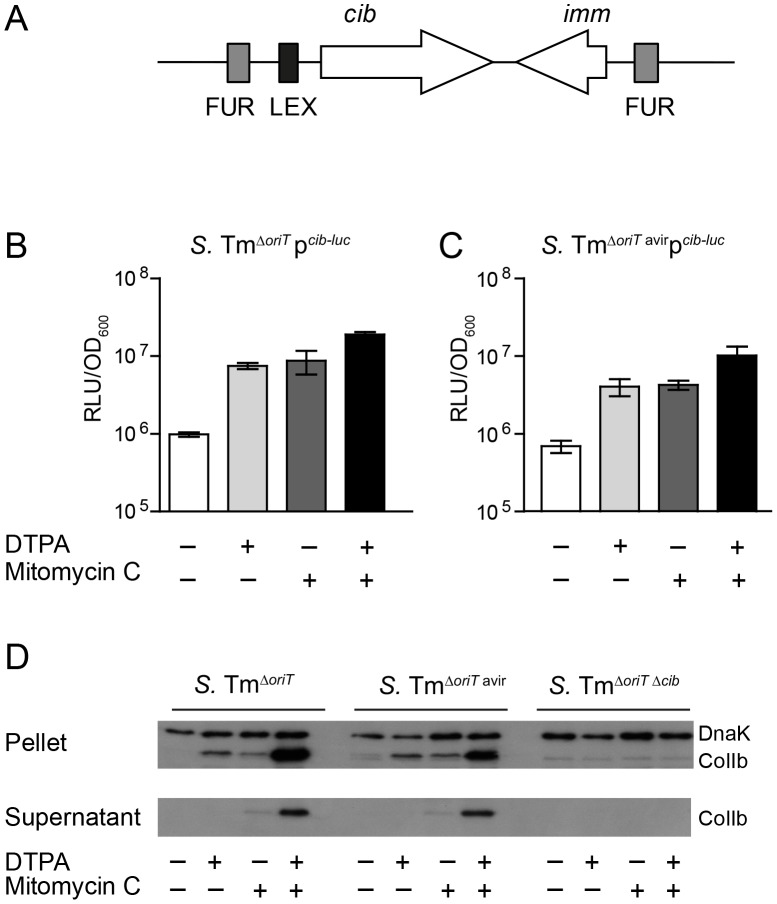
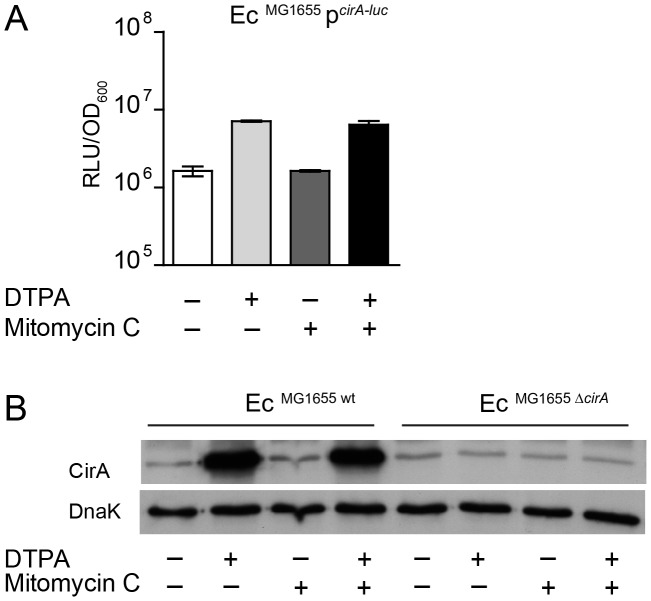
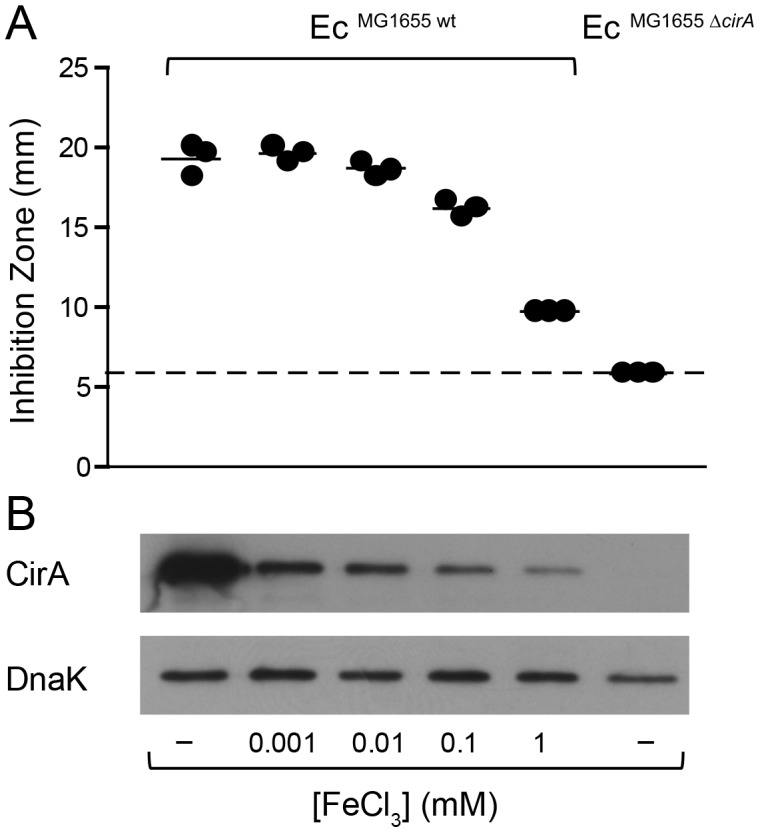
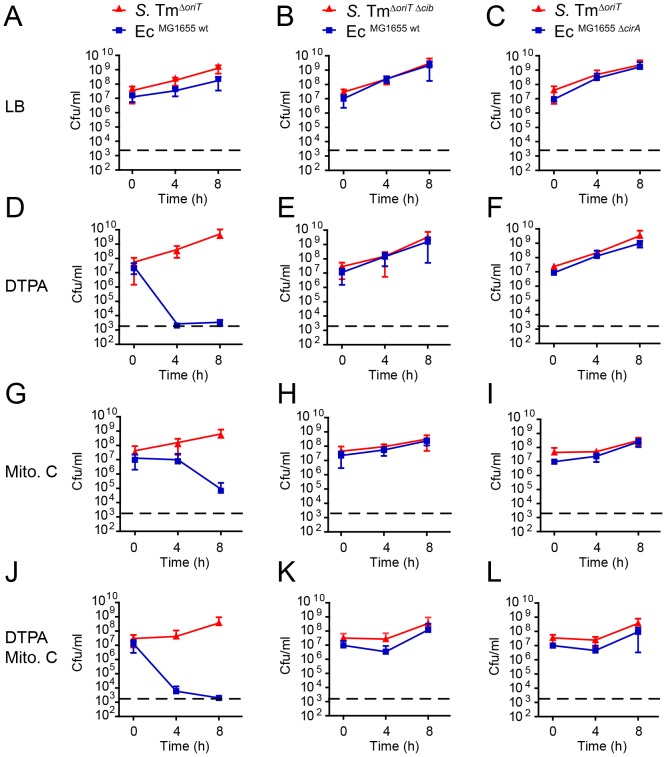
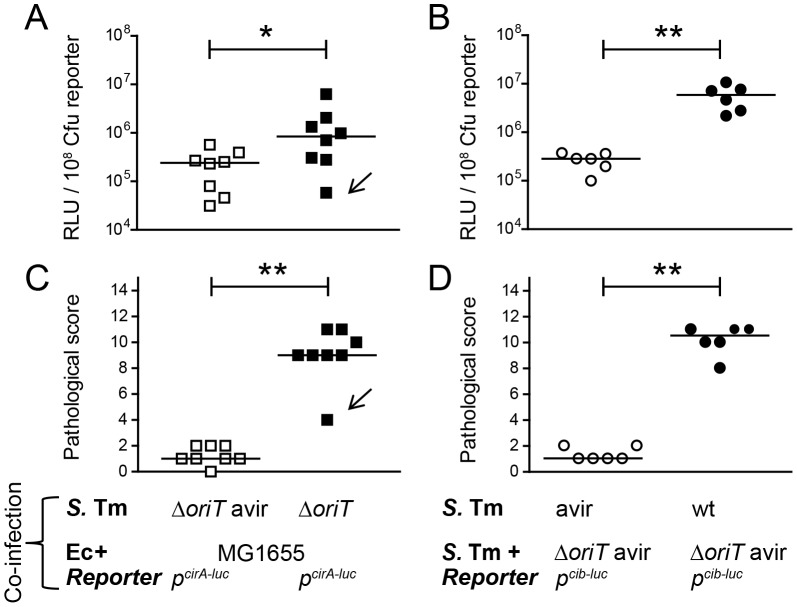
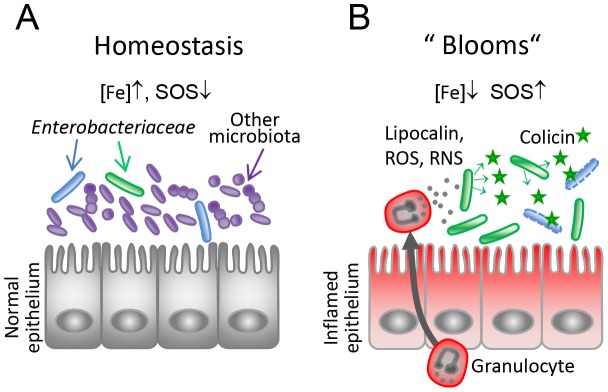
The host's immune system plays a key role in modulating growth of pathogens and the intestinal microbiota in the gut. In particular, inflammatory bowel disorders and pathogen infections induce shifts of the resident commensal microbiota which can result in overgrowth of Enterobacteriaceae ("inflammation-inflicted blooms"). Here, we investigated competition of the human pathogenic Salmonella enterica serovar Typhimurium strain SL1344 (S. Tm) and commensal E. coli in inflammation-inflicted blooms. S. Tm produces colicin Ib (ColIb), which is a narrow-spectrum protein toxin active against related Enterobacteriaceae. Production of ColIb conferred a competitive advantage to S. Tm over sensitive E. coli strains in the inflamed gut. In contrast, an avirulent S. Tm mutant strain defective in triggering gut inflammation did not benefit from ColIb. Expression of ColIb (cib) is regulated by iron limitation and the SOS response. CirA, the cognate outer membrane receptor of ColIb on colicin-sensitive E. coli, is induced upon iron limitation. We demonstrate that growth in inflammation-induced blooms favours expression of both S. Tm ColIb and the receptor CirA, thereby fuelling ColIb dependent competition of S. Tm and commensal E. coli in the gut. In conclusion, this study uncovers a so-far unappreciated role of inflammation-inflicted blooms as an environment favouring ColIb-dependent competition of pathogenic and commensal representatives of the Enterobacteriaceae family.
Conflict of interest statement
The authors declare no competing financial interests.
Figures
References
-
- Stecher B, Hardt WD (2010) Mechanisms controlling pathogen colonization of the gut. Curr Opin Microbiol 14 1: 82–91. - PubMed
-
- Sansonetti PJ (2008) Host-bacteria homeostasis in the healthy and inflamed gut. Curr Opin Gastroenterol 24: 435–439. - PubMed
-
- Lupp C, Robertson ML, Wickham ME, Sekirov I, Champion OL, et al. (2007) Host-mediated inflammation disrupts the intestinal microbiota and promotes the overgrowth of enterobacteriaceae. Cell, host and microbe 2: 119–129. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
