

Recruitment of PfSET2 by RNA polymerase II to variant antigen encoding loci contributes to antigenic variation in P. falciparum
- PMID: 24391504
- PMCID: PMC3879369
- DOI: 10.1371/journal.ppat.1003854
Recruitment of PfSET2 by RNA polymerase II to variant antigen encoding loci contributes to antigenic variation in P. falciparum
Abstract
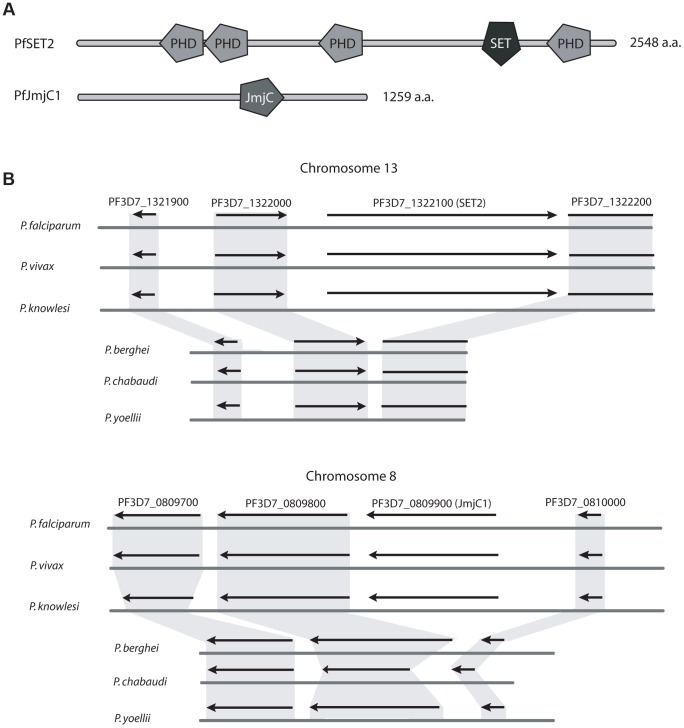
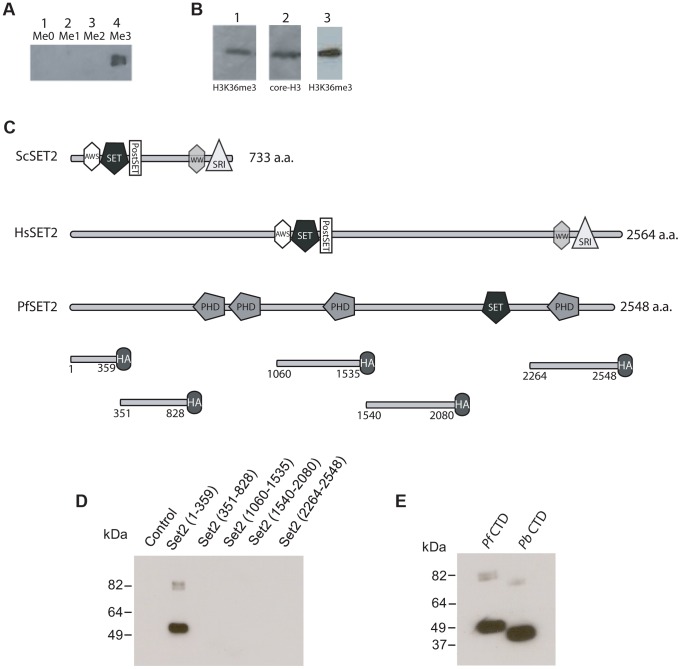
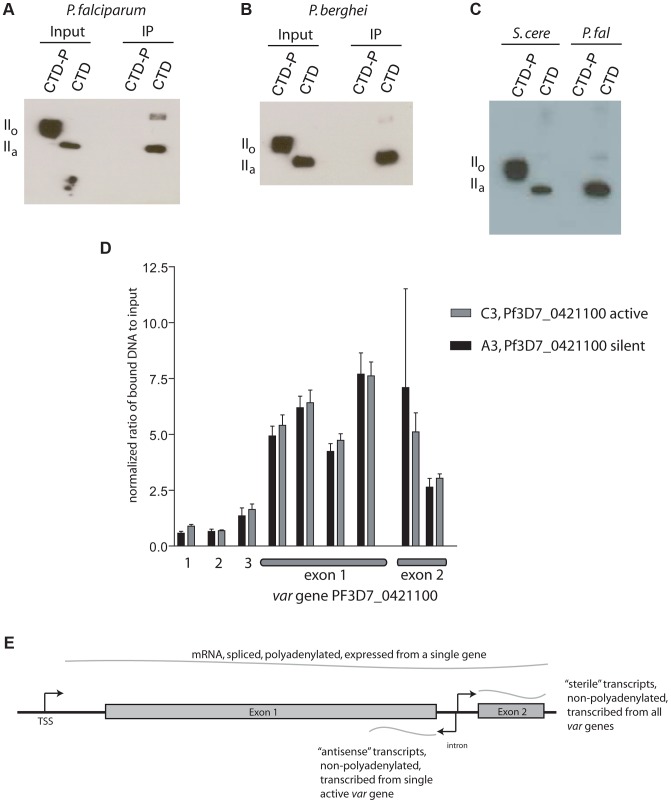
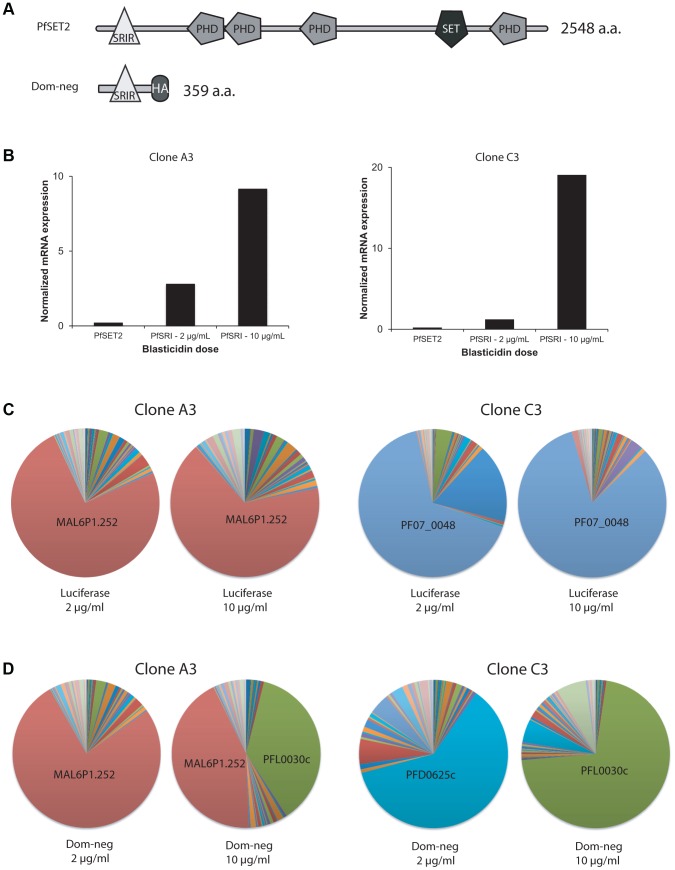
Histone modifications are important regulators of gene expression in all eukaryotes. In Plasmodium falciparum, these epigenetic marks regulate expression of genes involved in several aspects of host-parasite interactions, including antigenic variation. While the identities and genomic positions of many histone modifications have now been cataloged, how they are targeted to defined genomic regions remains poorly understood. For example, how variant antigen encoding loci (var) are targeted for deposition of unique histone marks is a mystery that continues to perplex the field. Here we describe the recruitment of an ortholog of the histone modifier SET2 to var genes through direct interactions with the C-terminal domain (CTD) of RNA polymerase II. In higher eukaryotes, SET2 is a histone methyltransferase recruited by RNA pol II during mRNA transcription; however, the ortholog in P. falciparum (PfSET2) has an atypical architecture and its role in regulating transcription is unknown. Here we show that PfSET2 binds to the unphosphorylated form of the CTD, a property inconsistent with its recruitment during mRNA synthesis. Further, we show that H3K36me3, the epigenetic mark deposited by PfSET2, is enriched at both active and silent var gene loci, providing additional evidence that its recruitment is not associated with mRNA production. Over-expression of a dominant negative form of PfSET2 designed to disrupt binding to RNA pol II induced rapid var gene expression switching, confirming both the importance of PfSET2 in var gene regulation and a role for RNA pol II in its recruitment. RNA pol II is known to transcribe non-coding RNAs from both active and silent var genes, providing a possible mechanism by which it could recruit PfSET2 to var loci. This work unifies previous reports of histone modifications, the production of ncRNAs, and the promoter activity of var introns into a mechanism that contributes to antigenic variation by malaria parasites.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
Similar articles
-
The Plasmodium falciparum histone methyltransferase PfSET10 is dispensable for the regulation of antigenic variation and gene expression in blood-stage parasites.mSphere. 2024 Nov 21;9(11):e0054624. doi: 10.1128/msphere.00546-24. Epub 2024 Oct 24. mSphere. 2024. PMID: 39445826 Free PMC article.
-
A Unique Virulence Gene Occupies a Principal Position in Immune Evasion by the Malaria Parasite Plasmodium falciparum.PLoS Genet. 2015 May 19;11(5):e1005234. doi: 10.1371/journal.pgen.1005234. eCollection 2015 May. PLoS Genet. 2015. PMID: 25993442 Free PMC article.
-
Plasmodium falciparum SET2 domain is allosterically regulated by its PHD-like domain to methylate at H3K36.Biochim Biophys Acta Gene Regul Mech. 2021 Oct;1864(10):194744. doi: 10.1016/j.bbagrm.2021.194744. Epub 2021 Aug 11. Biochim Biophys Acta Gene Regul Mech. 2021. PMID: 34389510
-
Antigenic variation in Plasmodium falciparum.Annu Rev Microbiol. 2008;62:445-70. doi: 10.1146/annurev.micro.61.080706.093134. Annu Rev Microbiol. 2008. PMID: 18785843 Review.
-
The epigenetic control of antigenic variation in Plasmodium falciparum.Curr Opin Microbiol. 2005 Aug;8(4):434-40. doi: 10.1016/j.mib.2005.06.007. Curr Opin Microbiol. 2005. PMID: 15979386 Review.
Cited by
-
Emerging biology of noncoding RNAs in malaria parasites.PLoS Pathog. 2022 Jul 7;18(7):e1010600. doi: 10.1371/journal.ppat.1010600. eCollection 2022 Jul. PLoS Pathog. 2022. PMID: 35797283 Free PMC article. Review.
-
In-depth comparative analysis of malaria parasite genomes reveals protein-coding genes linked to human disease in Plasmodium falciparum genome.BMC Genomics. 2018 May 2;19(1):312. doi: 10.1186/s12864-018-4654-5. BMC Genomics. 2018. PMID: 29716542 Free PMC article.
-
The role of long noncoding RNAs in malaria parasites.Trends Parasitol. 2023 Jul;39(7):517-531. doi: 10.1016/j.pt.2023.03.016. Epub 2023 Apr 29. Trends Parasitol. 2023. PMID: 37121862 Free PMC article. Review.
-
Disruption of the Plasmodium falciparum Life Cycle through Transcriptional Reprogramming by Inhibitors of Jumonji Demethylases.ACS Infect Dis. 2020 May 8;6(5):1058-1075. doi: 10.1021/acsinfecdis.9b00455. Epub 2020 Apr 24. ACS Infect Dis. 2020. PMID: 32272012 Free PMC article.
-
Recent advances in malaria genomics and epigenomics.Genome Med. 2016 Sep 7;8(1):92. doi: 10.1186/s13073-016-0343-7. Genome Med. 2016. PMID: 27605022 Free PMC article. Review.
References
-
- Crowley VM, Rovira-Graells N, Ribas dP, Cortes A (2011) Heterochromatin formation in bistable chromatin domains controls the epigenetic repression of clonally variant Plasmodium falciparum genes linked to erythrocyte invasion. Mol Microbiol 80: 391–406. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
