

NCYM, a Cis-antisense gene of MYCN, encodes a de novo evolved protein that inhibits GSK3β resulting in the stabilization of MYCN in human neuroblastomas
- PMID: 24391509
- PMCID: PMC3879166
- DOI: 10.1371/journal.pgen.1003996
NCYM, a Cis-antisense gene of MYCN, encodes a de novo evolved protein that inhibits GSK3β resulting in the stabilization of MYCN in human neuroblastomas
Abstract
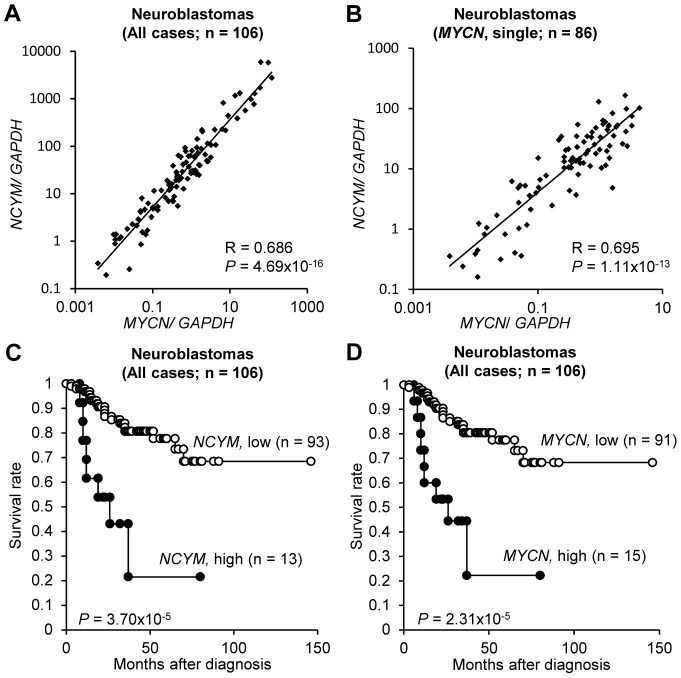
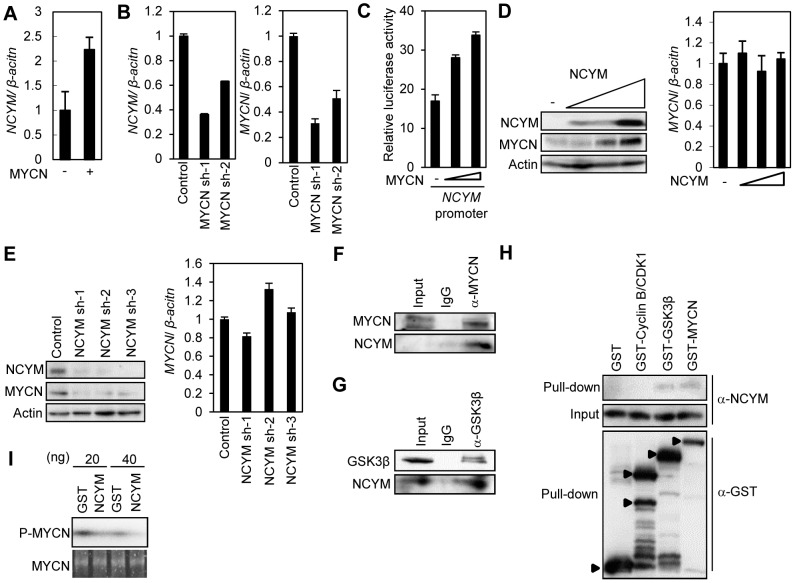
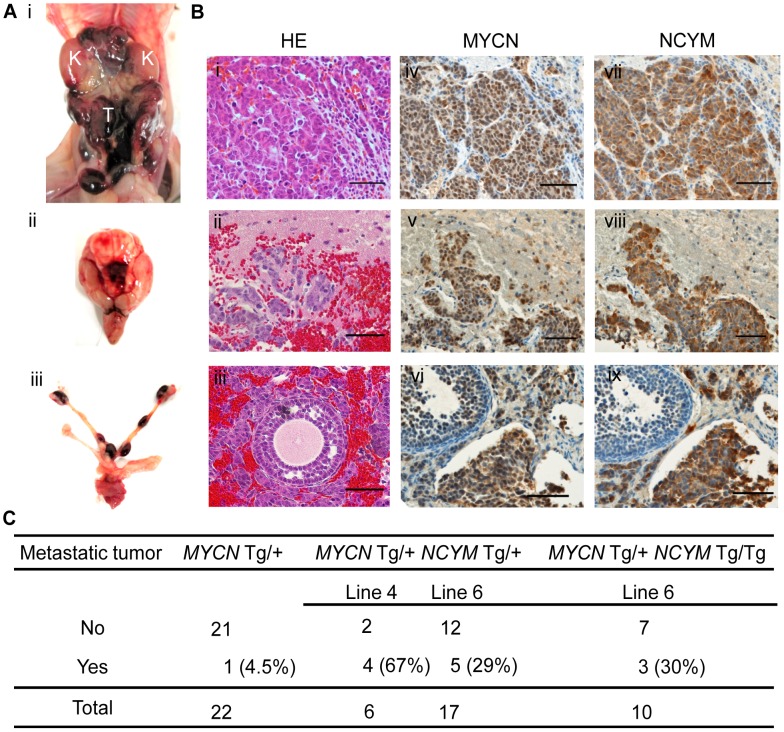
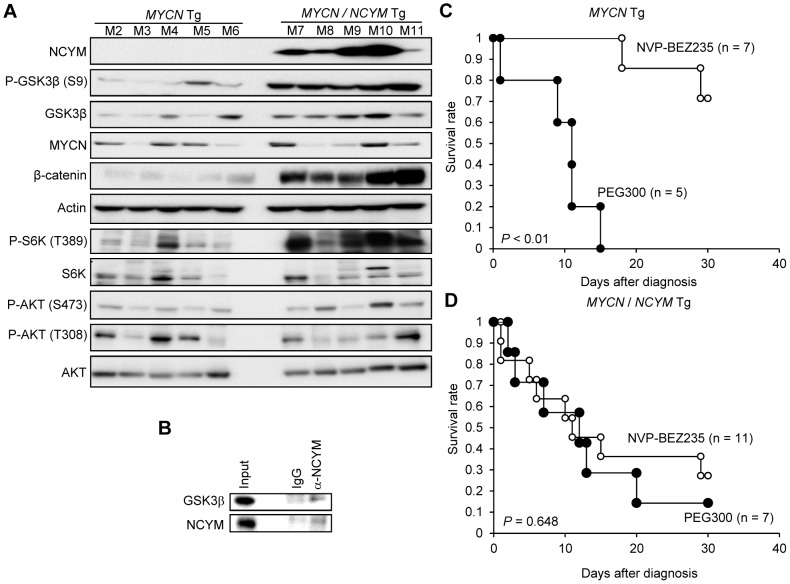
The rearrangement of pre-existing genes has long been thought of as the major mode of new gene generation. Recently, de novo gene birth from non-genic DNA was found to be an alternative mechanism to generate novel protein-coding genes. However, its functional role in human disease remains largely unknown. Here we show that NCYM, a cis-antisense gene of the MYCN oncogene, initially thought to be a large non-coding RNA, encodes a de novo evolved protein regulating the pathogenesis of human cancers, particularly neuroblastoma. The NCYM gene is evolutionally conserved only in the taxonomic group containing humans and chimpanzees. In primary human neuroblastomas, NCYM is 100% co-amplified and co-expressed with MYCN, and NCYM mRNA expression is associated with poor clinical outcome. MYCN directly transactivates both NCYM and MYCN mRNA, whereas NCYM stabilizes MYCN protein by inhibiting the activity of GSK3β, a kinase that promotes MYCN degradation. In contrast to MYCN transgenic mice, neuroblastomas in MYCN/NCYM double transgenic mice were frequently accompanied by distant metastases, behavior reminiscent of human neuroblastomas with MYCN amplification. The NCYM protein also interacts with GSK3β, thereby stabilizing the MYCN protein in the tumors of the MYCN/NCYM double transgenic mice. Thus, these results suggest that GSK3β inhibition by NCYM stabilizes the MYCN protein both in vitro and in vivo. Furthermore, the survival of MYCN transgenic mice bearing neuroblastoma was improved by treatment with NVP-BEZ235, a dual PI3K/mTOR inhibitor shown to destabilize MYCN via GSK3β activation. In contrast, tumors caused in MYCN/NCYM double transgenic mice showed chemo-resistance to the drug. Collectively, our results show that NCYM is the first de novo evolved protein known to act as an oncopromoting factor in human cancer, and suggest that de novo evolved proteins may functionally characterize human disease.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures

References
-
- Jacob F (1977) Evolution and tinkering. Science 196: 1161–1166. - PubMed
-
- Ohno S (1970) Evolution by gene duplication. New York, NY, Springer-Verlag.
-
- Tautz D, Domazet-Loso T (2011) The evolutionary origin of orphan genes. Nat Rev Genet 12: 692–702. - PubMed
-
- Khalturin K, Hemmrich G, Fraune S, Augustin R, Bosch TC (2009) More than just orphans: are taxonomically-restricted genes important in evolution? Trends Genet 25: 404–413. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
