

Root-expressed maize lipoxygenase 3 negatively regulates induced systemic resistance to Colletotrichum graminicola in shoots
- PMID: 24391653
- PMCID: PMC3867115
- DOI: 10.3389/fpls.2013.00510
Root-expressed maize lipoxygenase 3 negatively regulates induced systemic resistance to Colletotrichum graminicola in shoots
Abstract
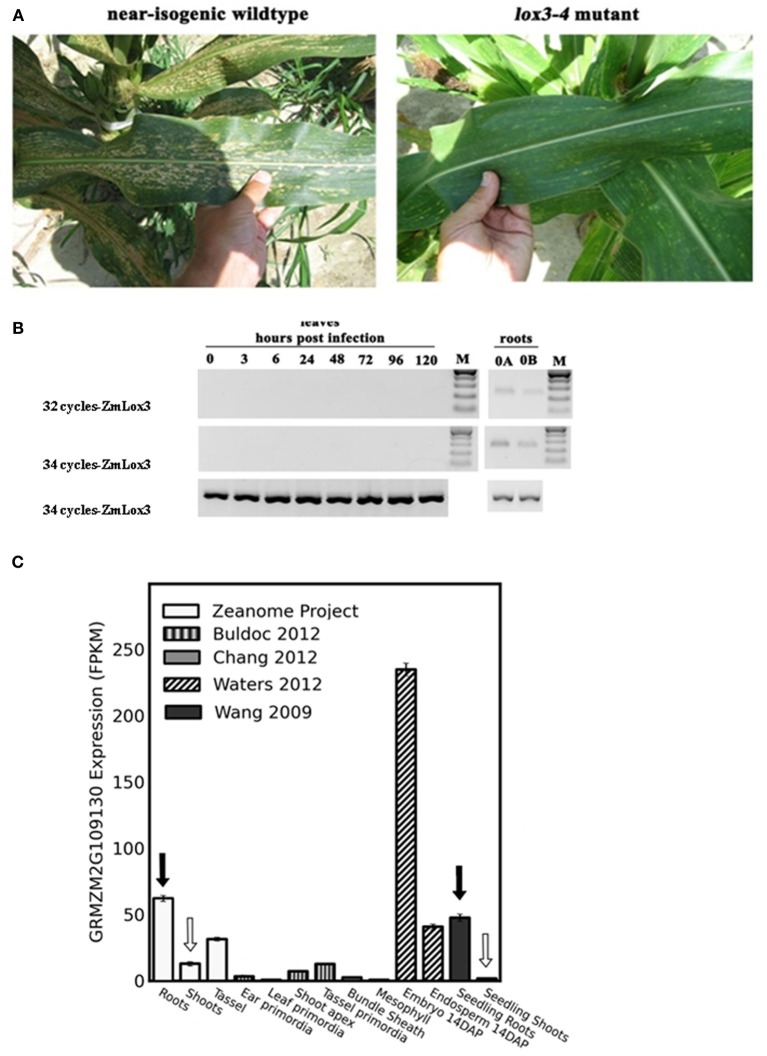
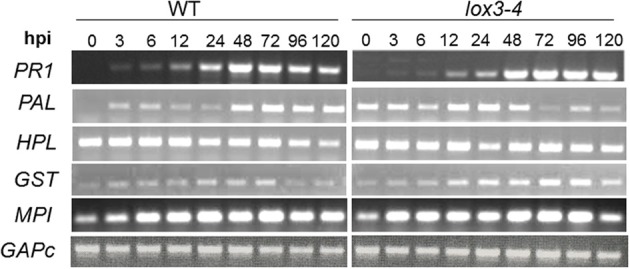
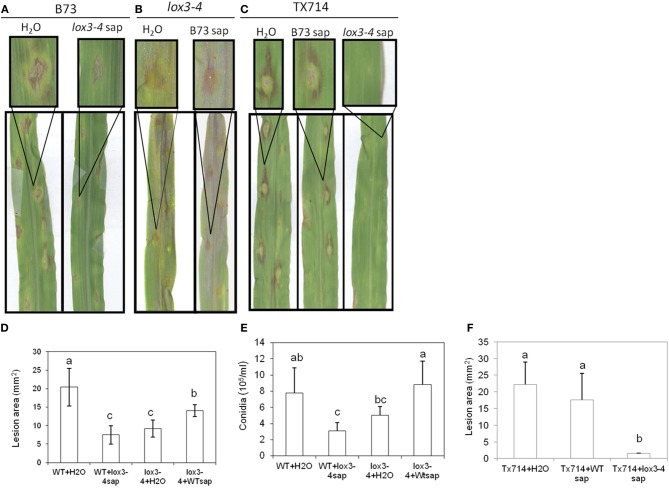
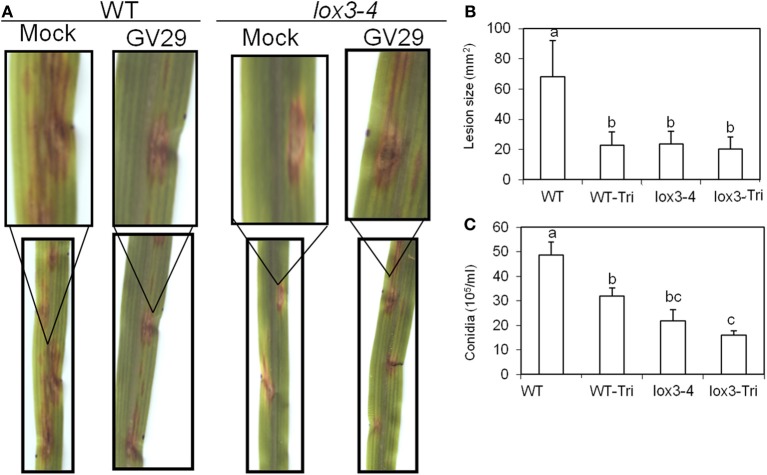
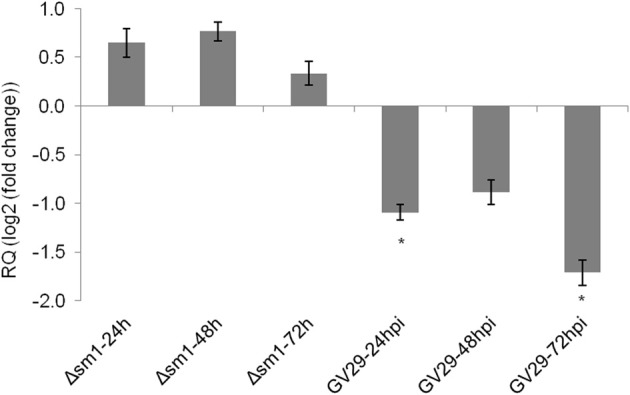
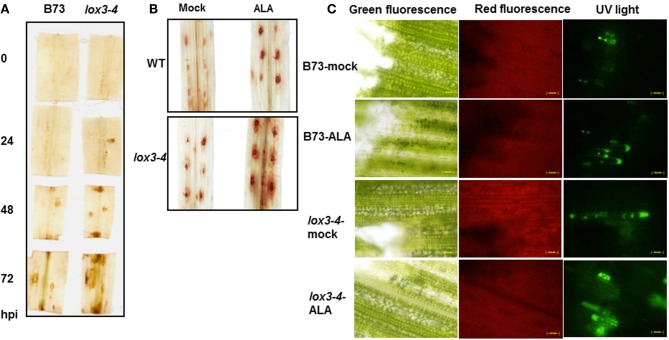
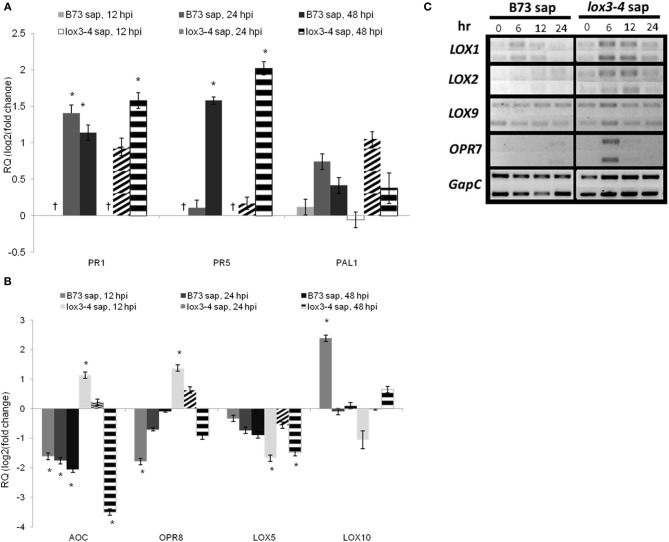
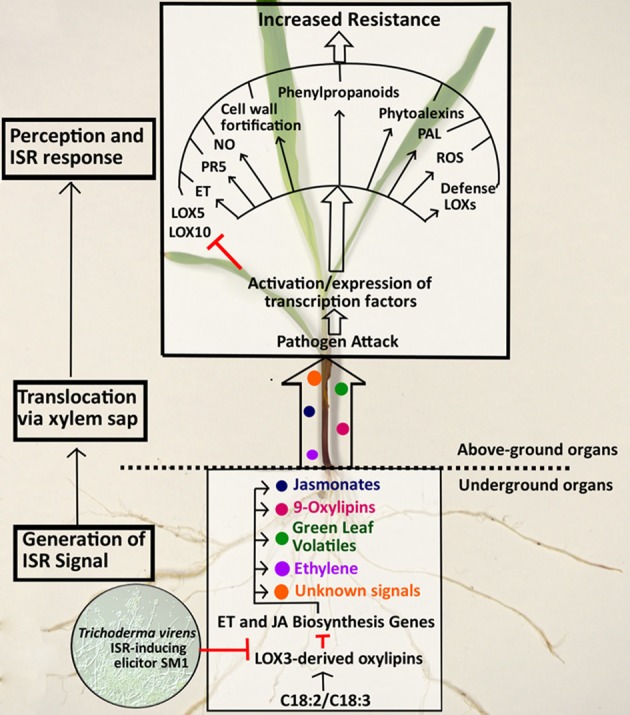
We have previously reported that disruption of a maize root-expressed 9-lipoxygenase (9-LOX) gene, ZmLOX3, results in dramatic increase in resistance to diverse leaf and stalk pathogens. Despite evident economic significance of these findings, the mechanism behind this increased resistance remained elusive. In this study, we found that increased resistance of the lox3-4 mutants is due to constitutive activation of induced systemic resistance (ISR) signaling. We showed that ZmLOX3 lacked expression in leaves in response to anthracnose leaf blight pathogen Colletotrichum graminicola, but was expressed constitutively in the roots, thus, prompting our hypothesis: the roots of lox3-4 mutants are the source of increased resistance in leaves. Supporting this hypothesis, treatment of wild-type plants (WT) with xylem sap of lox3-4 mutant induced resistance to C. graminicola to the levels comparable to those observed in lox3-4 mutant. Moreover, treating mutants with the sap collected from WT plants partially restored the susceptibility to C. graminicola. lox3-4 mutants showed primed defense responses upon infection, which included earlier and greater induction of defense-related PAL and GST genes compared to WT. In addition to the greater expression of the octadecanoid pathway genes, lox3-4 mutant responded earlier and with a greater accumulation of H2O2 in response to C. graminicola infection or treatment with alamethicin. These findings suggest that lox3-4 mutants display constitutive ISR-like signaling. In support of this idea, root colonization by Trichoderma virens strain GV29-8 induced the same level of disease resistance in WT as the treatment with the mutant sap, but had no additional resistance effect in lox3-4 mutant. While treatment with T. virens GV29 strongly and rapidly suppressed ZmLOX3 expression in hydroponically grown WT roots, T. virens Δsml mutant, which is deficient in ISR induction, was unable to suppress expression of ZmLOX3, thus, providing genetic evidence that SM1 function in ISR, at least in part, by suppressing host ZmLOX3 gene. This study and the genetic tools generated herein will allow the identification of the signals regulating the induction of resistance to aboveground attackers by beneficial soil microorganisms in the future.
Keywords: Colletotrichum graminicola; Trichoderma; beneficial microorganisms; hydrogen peroxide; long distance signaling; oxylipin; priming; root-to-shoot signaling.
Figures
References
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous