

Base preferences in non-templated nucleotide incorporation by MMLV-derived reverse transcriptases
- PMID: 24392002
- PMCID: PMC3877366
- DOI: 10.1371/journal.pone.0085270
Base preferences in non-templated nucleotide incorporation by MMLV-derived reverse transcriptases
Abstract
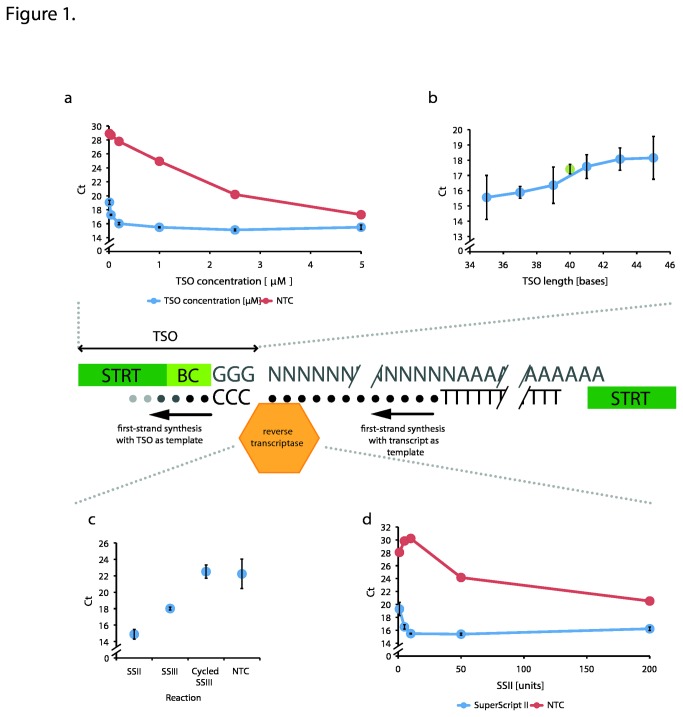
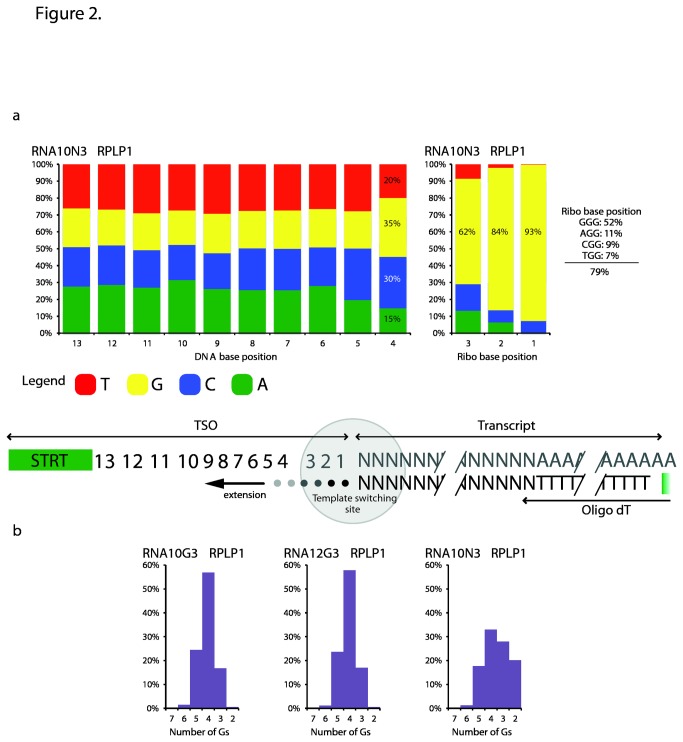
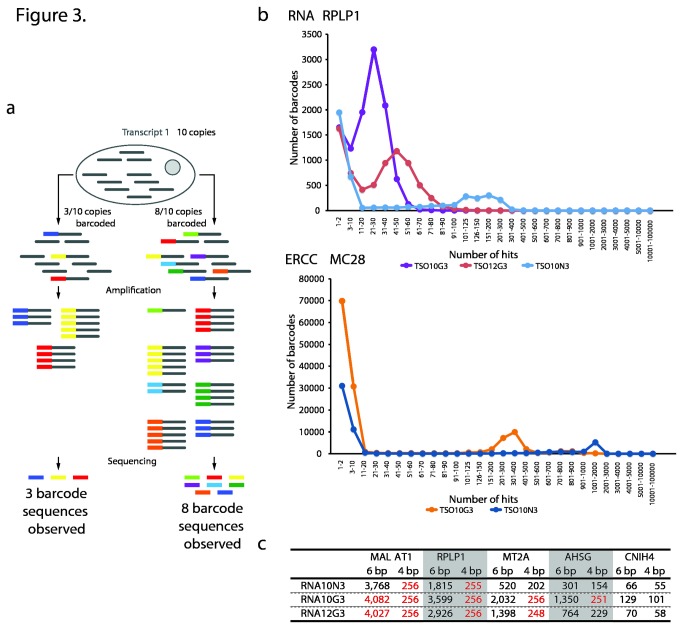
Reverse transcriptases derived from Moloney Murine Leukemia Virus (MMLV) have an intrinsic terminal transferase activity, which causes the addition of a few non-templated nucleotides at the 3' end of cDNA, with a preference for cytosine. This mechanism can be exploited to make the reverse transcriptase switch template from the RNA molecule to a secondary oligonucleotide during first-strand cDNA synthesis, and thereby to introduce arbitrary barcode or adaptor sequences in the cDNA. Because the mechanism is relatively efficient and occurs in a single reaction, it has recently found use in several protocols for single-cell RNA sequencing. However, the base preference of the terminal transferase activity is not known in detail, which may lead to inefficiencies in template switching when starting from tiny amounts of mRNA. Here, we used fully degenerate oligos to determine the exact base preference at the template switching site up to a distance of ten nucleotides. We found a strong preference for guanosine at the first non-templated nucleotide, with a greatly reduced bias at progressively more distant positions. Based on this result, and a number of careful optimizations, we report conditions for efficient template switching for cDNA amplification from single cells.
Conflict of interest statement
Figures
Similar articles
-
Non-templated addition and template switching by Moloney murine leukemia virus (MMLV)-based reverse transcriptases co-occur and compete with each other.J Biol Chem. 2019 Nov 29;294(48):18220-18231. doi: 10.1074/jbc.RA119.010676. Epub 2019 Oct 22. J Biol Chem. 2019. PMID: 31640989 Free PMC article.
-
RNA-dependent DNA polymerases.Curr Protoc Mol Biol. 2008 Oct;Chapter 3:Unit3.7. doi: 10.1002/0471142727.mb0307s84. Curr Protoc Mol Biol. 2008. PMID: 18972389
-
Crystal structures of an N-terminal fragment from Moloney murine leukemia virus reverse transcriptase complexed with nucleic acid: functional implications for template-primer binding to the fingers domain.J Mol Biol. 2000 Feb 18;296(2):613-32. doi: 10.1006/jmbi.1999.3477. J Mol Biol. 2000. PMID: 10669612
-
Translesion synthesis by AMV, HIV, and MMLVreverse transcriptases using RNA templates containing inosine, guanosine, and their 8-oxo-7,8-dihydropurine derivatives.PLoS One. 2020 Aug 28;15(8):e0235102. doi: 10.1371/journal.pone.0235102. eCollection 2020. PLoS One. 2020. PMID: 32857764 Free PMC article.
-
[Inhibition of gene expression by antisense DNA].Nihon Rinsho. 1995 Mar;53(3):771-8. Nihon Rinsho. 1995. PMID: 7535367 Review. Japanese.
Cited by
-
NanoCAGE-XL and CapFilter: an approach to genome wide identification of high confidence transcription start sites.BMC Genomics. 2015 Aug 13;16(1):597. doi: 10.1186/s12864-015-1670-6. BMC Genomics. 2015. PMID: 26268438 Free PMC article.
-
FB5P-seq: FACS-Based 5-Prime End Single-Cell RNA-seq for Integrative Analysis of Transcriptome and Antigen Receptor Repertoire in B and T Cells.Front Immunol. 2020 Mar 3;11:216. doi: 10.3389/fimmu.2020.00216. eCollection 2020. Front Immunol. 2020. PMID: 32194545 Free PMC article.
-
Compounds that enhance the tailing activity of Moloney murine leukemia virus reverse transcriptase.Sci Rep. 2017 Jul 26;7(1):6520. doi: 10.1038/s41598-017-04765-8. Sci Rep. 2017. PMID: 28747695 Free PMC article.
-
CRISPR with Transcriptional Readout reveals influenza transcription is modulated by NELF and can precipitate an interferon response.bioRxiv [Preprint]. 2025 Jun 17:2024.11.14.623683. doi: 10.1101/2024.11.14.623683. bioRxiv. 2025. PMID: 39605461 Free PMC article. Preprint.
-
Methodologies for Transcript Profiling Using Long-Read Technologies.Front Genet. 2020 Jul 7;11:606. doi: 10.3389/fgene.2020.00606. eCollection 2020. Front Genet. 2020. PMID: 32733532 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
