

Determinants for the improved thermostability of a mesophilic family 11 xylanase predicted by computational methods
- PMID: 24393334
- PMCID: PMC3895927
- DOI: 10.1186/1754-6834-7-3
Determinants for the improved thermostability of a mesophilic family 11 xylanase predicted by computational methods
Abstract
Background: Xylanases have drawn much attention owing to possessing great potential in various industrial applications. However, the applicability of xylanases, exemplified by the production of bioethanol and xylooligosaccharides (XOSs), was bottlenecked by their low stabilities at higher temperatures. The main purpose of this work was to improve the thermostability of AuXyn11A, a mesophilic glycoside hydrolase (GH) family 11 xylanase from Aspergillus usamii E001, by N-terminus replacement.
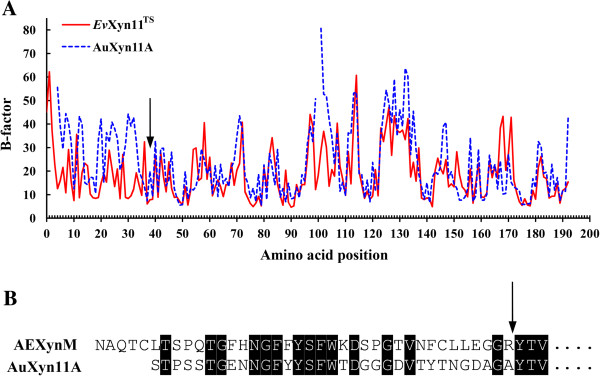
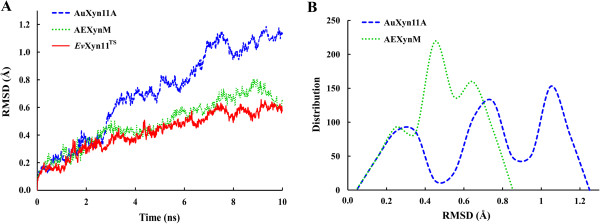
Results: A hybrid xylanase with high thermostability, named AEXynM, was predicted by computational methods, and constructed by substituting the N-terminal 33 amino acids of AuXyn11A with the corresponding 38 ones of EvXyn11TS, a hyperthermostable family 11 xylanase. Two AuXyn11A- and AEXynM-encoding genes, Auxyn11A and AExynM, were then highly expressed in Pichia pastoris GS115, respectively. The specific activities of two recombinant xylanases (reAuXyn11A and reAEXynM) were 10,437 and 9,529 U mg-1. The temperature optimum and stability of reAEXynM reached 70 and 75°C, respectively, much higher than those (50 and 45°C) of reAuXyn11A. The melting temperature (Tm) of reAEXynM, measured using the Protein Thermal Shift (PTS) method, increased by 34.0°C as compared with that of reAuXyn11A. Analyzed by HPLC, xylobiose and xylotriose as the major hydrolytic products were excised from corncob xylan by reAEXynM. Additionally, three single mutant genes from AExynM (AExynMC5T, AExynMP9S, and AExynMH14N) were constructed by site-directed mutagenesis as designed theoretically, and expressed in P. pastoris GS115, respectively. The thermostabilities of three recombinant mutants clearly decreased as compared with that of reAEXynM, which demonstrated that the three amino acids (Cys5, Pro9, and His14) in the replaced N-terminus contributed mainly to the high thermostability of AEXynM.
Conclusions: This work highly enhanced the thermostability of AuXyn11A by N-terminus replacement, and further verified, by site-directed mutagenesis, that Cys5, Pro9, and His14 contributed mainly to the improved thermostability. It will provide an effective strategy for improving the thermostabilities of other enzymes.
Figures





Similar articles
-
Engineering mesophilic GH11 xylanase from Cellulomonas flavigena by rational design of N-terminus substitution.Front Bioeng Biotechnol. 2022 Nov 3;10:1044291. doi: 10.3389/fbioe.2022.1044291. eCollection 2022. Front Bioeng Biotechnol. 2022. PMID: 36406235 Free PMC article.
-
Optimized expression, purification and characterization of a family 11 xylanase (AuXyn11A) from Aspergillus usamii E001 in Pichia pastoris.J Sci Food Agric. 2014 Mar 15;94(4):699-706. doi: 10.1002/jsfa.6309. Epub 2013 Aug 5. J Sci Food Agric. 2014. PMID: 23881861
-
Enhanced thermostability of a mesophilic xylanase by N-terminal replacement designed by molecular dynamics simulation.J Sci Food Agric. 2013 Sep;93(12):3016-23. doi: 10.1002/jsfa.6134. Epub 2013 Apr 19. J Sci Food Agric. 2013. PMID: 23512640
-
[Effect of N-terminal disulfide bridge on thermostability of family 11 xylanases].Sheng Wu Gong Cheng Xue Bao. 2012 Dec;28(12):1441-9. Sheng Wu Gong Cheng Xue Bao. 2012. PMID: 23593868 Chinese.
-
[Correlation between thermostability of the xylanase EvXyn11(TS) and its N-terminal disulfide bridge].Wei Sheng Wu Xue Bao. 2013 Apr 4;53(4):346-52. Wei Sheng Wu Xue Bao. 2013. PMID: 23858709 Chinese.
Cited by
-
Directed Modification of a GHF11 Thermostable Xylanase AusM for Enhancing Inhibitory Resistance towards SyXIP-I and Application of AusMPKK in Bread Making.Foods. 2023 Sep 26;12(19):3574. doi: 10.3390/foods12193574. Foods. 2023. PMID: 37835228 Free PMC article.
-
Engineering mesophilic GH11 xylanase from Cellulomonas flavigena by rational design of N-terminus substitution.Front Bioeng Biotechnol. 2022 Nov 3;10:1044291. doi: 10.3389/fbioe.2022.1044291. eCollection 2022. Front Bioeng Biotechnol. 2022. PMID: 36406235 Free PMC article.
-
Design, expression and functional characterization of a thermostable xylanase from Trichoderma reesei.PLoS One. 2019 Jan 16;14(1):e0210548. doi: 10.1371/journal.pone.0210548. eCollection 2019. PLoS One. 2019. PMID: 30650138 Free PMC article.
-
An Age Effect of Rumen Microbiome in Dairy Buffaloes Revealed by Metagenomics.Microorganisms. 2022 Jul 25;10(8):1491. doi: 10.3390/microorganisms10081491. Microorganisms. 2022. PMID: 35893549 Free PMC article.
-
Improving the thermostability of a fungal GH11 xylanase via site-directed mutagenesis guided by sequence and structural analysis.Biotechnol Biofuels. 2017 May 23;10:133. doi: 10.1186/s13068-017-0824-y. eCollection 2017. Biotechnol Biofuels. 2017. PMID: 28546828 Free PMC article.
References
-
- Taibi Z, Saoudi B, Boudelaa M, Trigui H, Belghith H, Gargouri A, Ladjama A. Purification and biochemical characterization of a highly thermostable xylanase from Actinomadura sp. strain Cpt20 isolated from poultry compost. Appl Biochem Biotechnol. 2012;166(3):663–679. doi: 10.1007/s12010-011-9457-y. - DOI - PubMed
-
- Zhang J, Siika-aho M, Puranen T, Tang M, Tenkanen M, Viikari L. Thermostable recombinant xylanases from Nonomuraea flexuosa and Thermoascus aurantiacus show distinct properties in the hydrolysis of xylans and pretreated wheat straw. Biotechnol Biofuels. 2011;4:12. doi: 10.1186/1754-6834-4-12. - DOI - PMC - PubMed
LinkOut - more resources
Full Text Sources
Other Literature Sources
