

Mammalian conserved ADAR targets comprise only a small fragment of the human editosome
- PMID: 24393560
- PMCID: PMC4053846
- DOI: 10.1186/gb-2014-15-1-r5
Mammalian conserved ADAR targets comprise only a small fragment of the human editosome
Abstract
Background: ADAR proteins are among the most extensively studied RNA binding proteins. They bind to their target and deaminate specific adenosines to inosines. ADAR activity is essential, and the editing of a subset of their targets is critical for viability. Recently, a huge number of novel ADAR targets were detected by analyzing next generation sequencing data. Most of these novel editing sites are located in lineage-specific genomic repeats, probably a result of overactivity of editing enzymes, thus masking the functional sites. In this study we aim to identify the set of mammalian conserved ADAR targets.
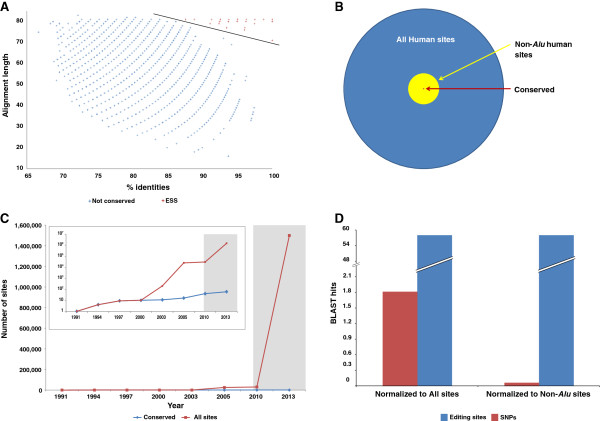
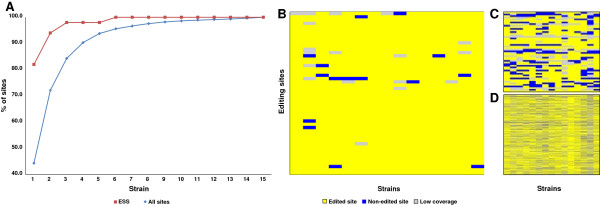
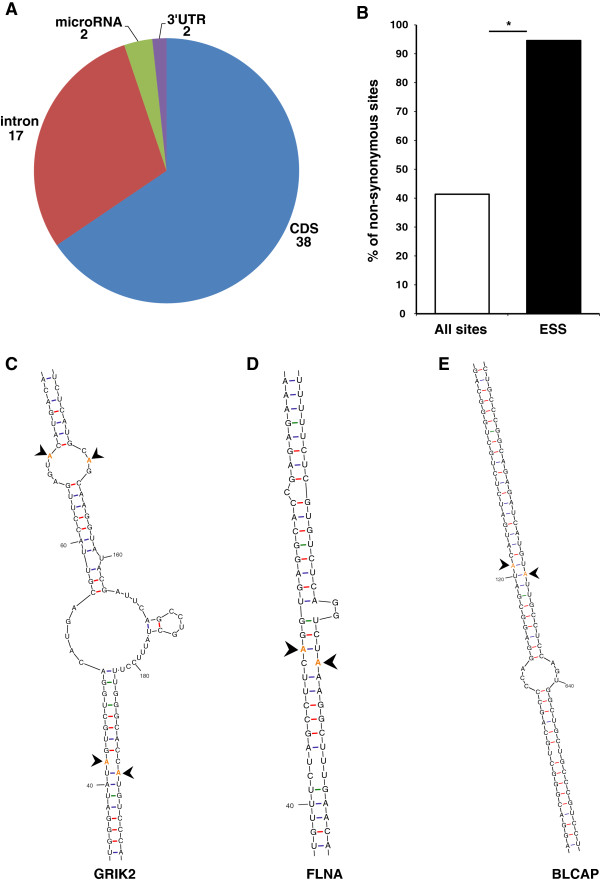
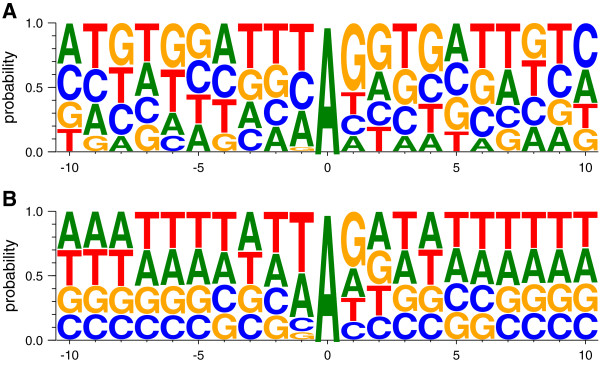
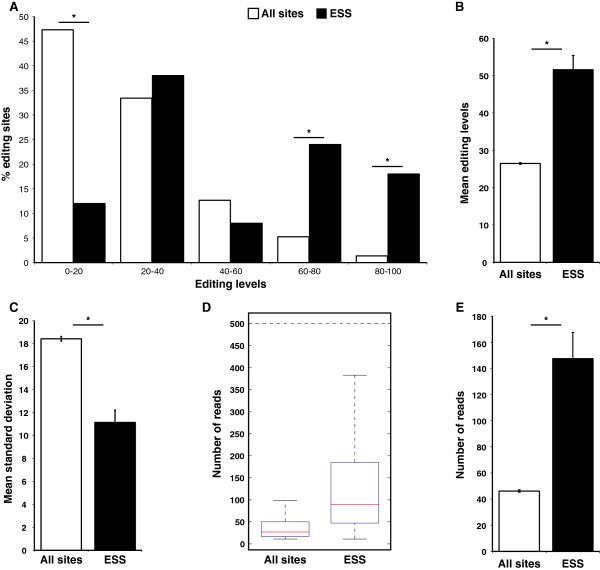
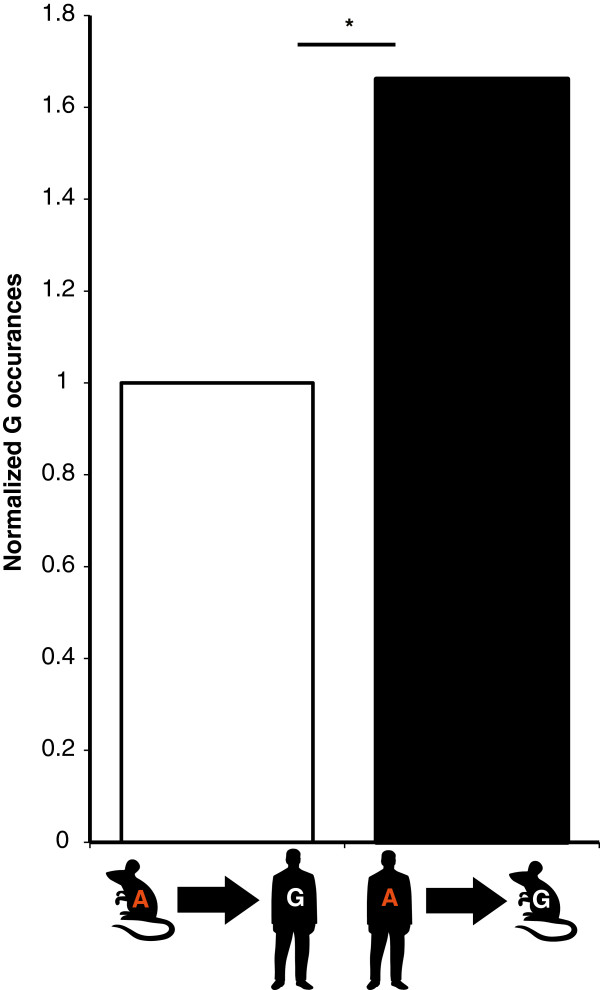
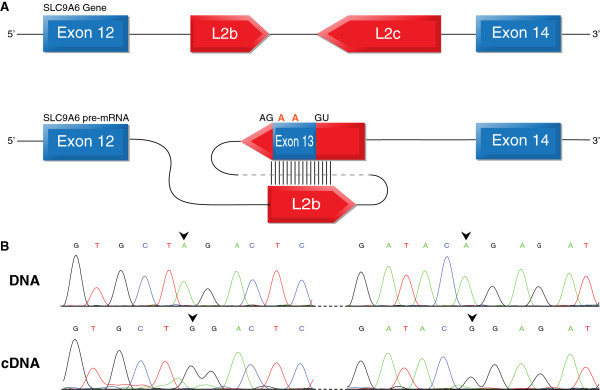
Results: We used RNA sequencing data from human, mouse, rat, cow, opossum, and platypus to define the conserved mammalian set of ADAR targets. We found that the conserved mammalian editing sites are surprisingly small in number and have unique characteristics that distinguish them from non-conserved ones. The sites that constitute the set have a distinct genomic distribution, tend to be located in genes encoding neurotransmitter receptors or other synapse related proteins, and have higher editing and expression levels. We also found a high consistency of editing levels of this set within mice strains and between human and mouse. Tight regulation of editing in these sites across strains and species implies their functional importance.
Conclusions: Despite the discovery of numerous editing targets, only a small number of them are conserved within mammalian evolution. These sites are extremely highly conserved and exhibit unique features, such as tight regulation, and probably play a pivotal role in mammalian biology.
Figures
Comment in
-
Identification of evolutionarily meaningful information within the mammalian RNA editing landscape.Genome Biol. 2014 Jan 28;15(1):103. doi: 10.1186/gb4157. Genome Biol. 2014. PMID: 24468094 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
