

Ankyrin-B regulates Cav2.1 and Cav2.2 channel expression and targeting
- PMID: 24394417
- PMCID: PMC3931084
- DOI: 10.1074/jbc.M113.523639
Ankyrin-B regulates Cav2.1 and Cav2.2 channel expression and targeting
Abstract
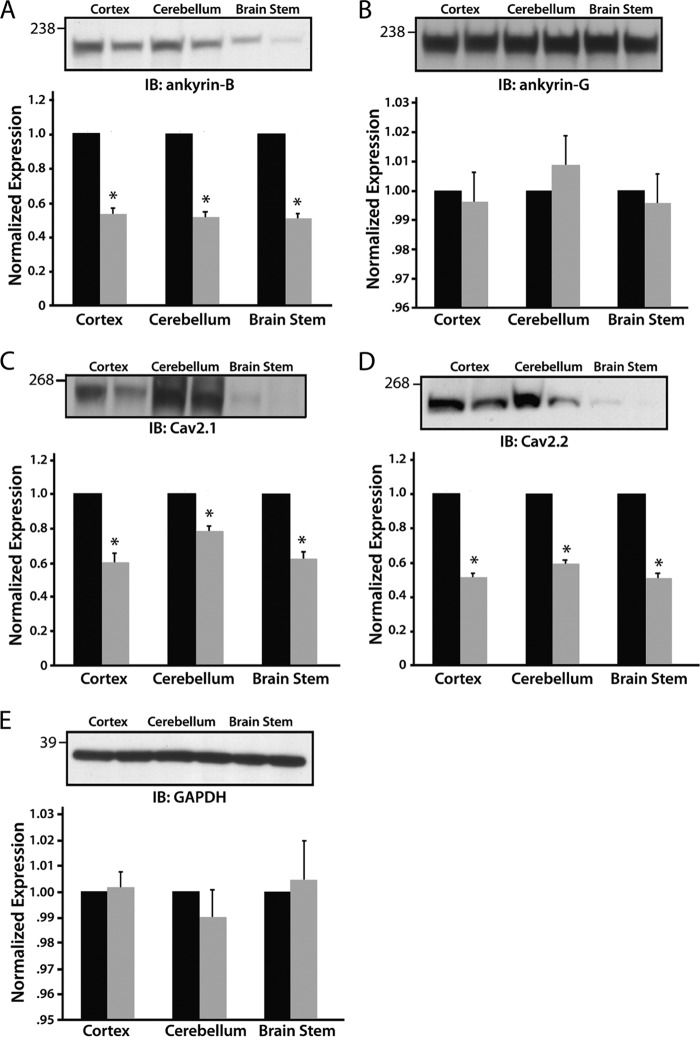
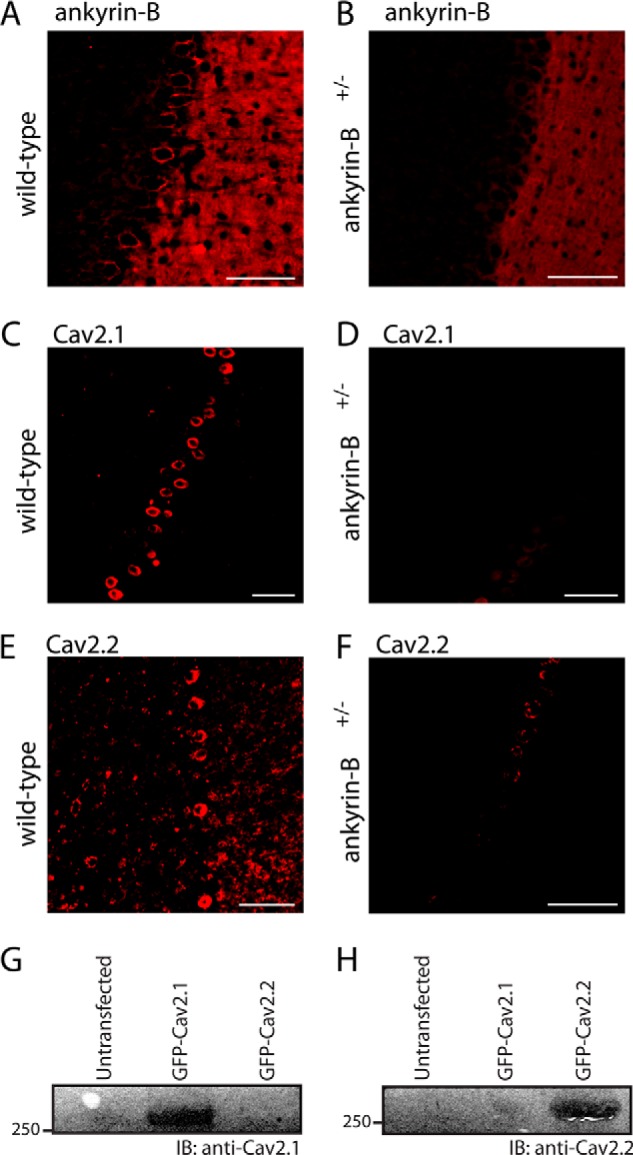
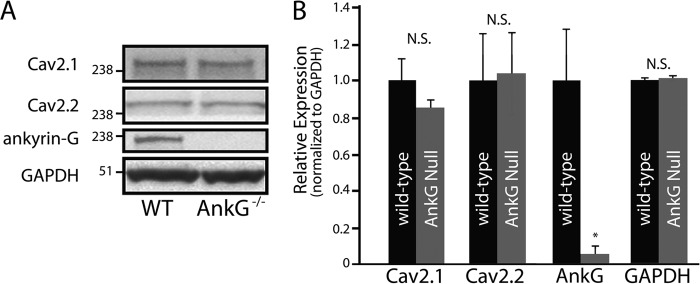
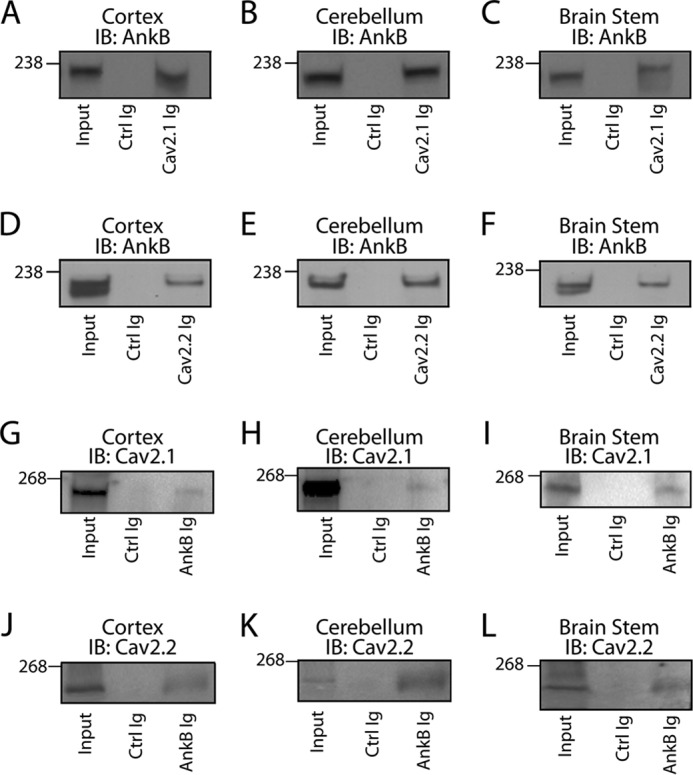
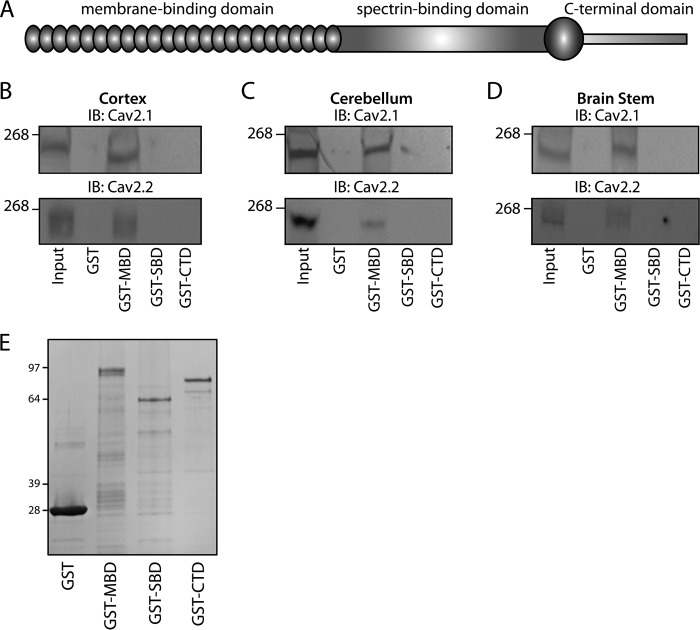
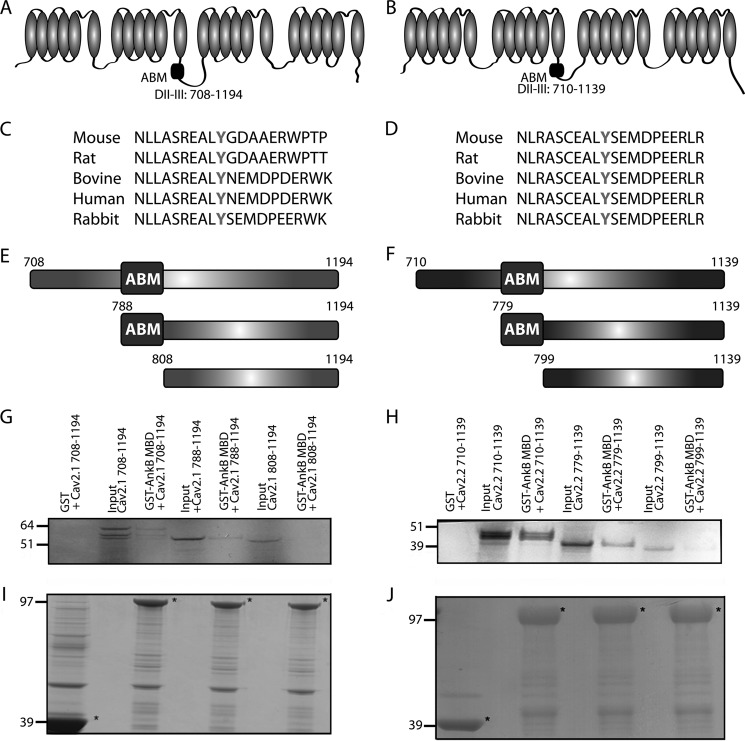
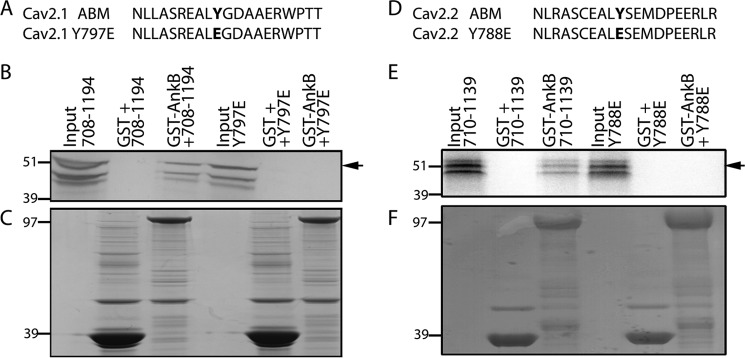
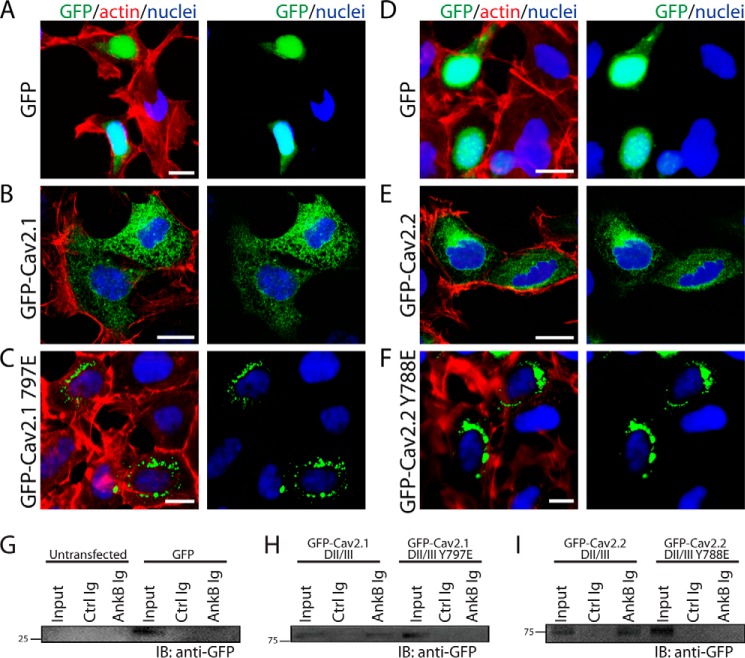
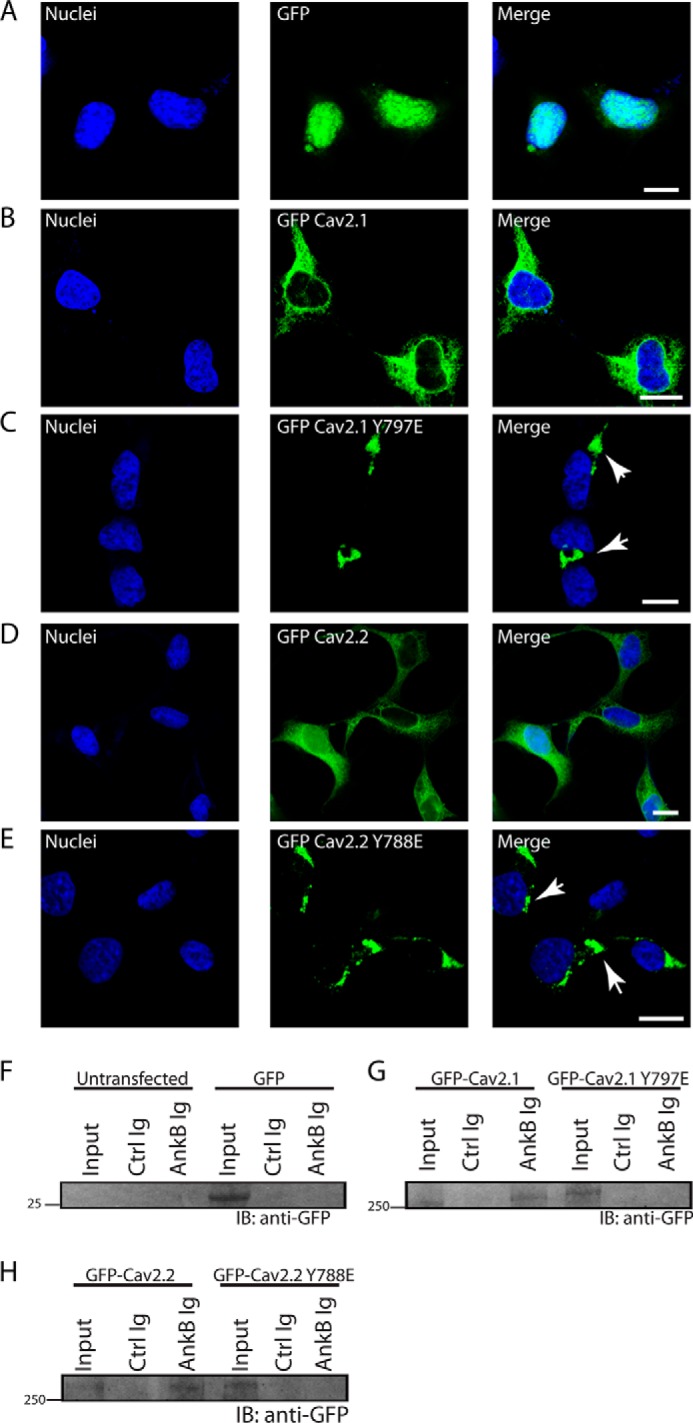
N-type and P/Q-type calcium channels are documented players in the regulation of synaptic function; however, the mechanisms underlying their expression and cellular targeting are poorly understood. Ankyrin polypeptides are essential for normal integral membrane protein expression in a number of cell types, including neurons, cardiomyocytes, epithelia, secretory cells, and erythrocytes. Ankyrin dysfunction has been linked to defects in integral protein expression, abnormal cellular function, and disease. Here, we demonstrate that ankyrin-B associates with Cav2.1 and Cav2.2 in cortex, cerebellum, and brain stem. Additionally, using in vitro and in vivo techniques, we demonstrate that ankyrin-B, via its membrane-binding domain, associates with a highly conserved motif in the DII/III loop domain of Cav2.1 and Cav2.2. Further, we demonstrate that this domain is necessary for proper targeting of Cav2.1 and Cav2.2 in a heterologous system. Finally, we demonstrate that mutation of a single conserved tyrosine residue in the ankyrin-binding motif of both Cav2.1 (Y797E) and Cav2.2 (Y788E) results in loss of association with ankyrin-B in vitro and in vivo. Collectively, our findings identify an interaction between ankyrin-B and both Cav2.1 and Cav2.2 at the amino acid level that is necessary for proper Cav2.1 and Cav2.2 targeting in vivo.
Keywords: Adaptor Proteins; Cytoskeleton; Ion Channels; Membrane Trafficking; Protein Sorting; Protein Targeting.
Figures
Similar articles
-
Ankyrin B and Ankyrin B variants differentially modulate intracellular and surface Cav2.1 levels.Mol Brain. 2019 Sep 2;12(1):75. doi: 10.1186/s13041-019-0494-8. Mol Brain. 2019. PMID: 31477143 Free PMC article.
-
Alternative splicing generates a smaller assortment of CaV2.1 transcripts in cerebellar Purkinje cells than in the cerebellum.Physiol Genomics. 2006 Jan 12;24(2):86-96. doi: 10.1152/physiolgenomics.00149.2005. Epub 2005 Nov 8. Physiol Genomics. 2006. PMID: 16278278
-
Obscurin targets ankyrin-B and protein phosphatase 2A to the cardiac M-line.J Biol Chem. 2008 Nov 14;283(46):31968-80. doi: 10.1074/jbc.M806050200. Epub 2008 Sep 9. J Biol Chem. 2008. PMID: 18782775 Free PMC article.
-
Cardiac ankyrins: Essential components for development and maintenance of excitable membrane domains in heart.Cardiovasc Res. 2006 Jul 1;71(1):22-9. doi: 10.1016/j.cardiores.2006.03.018. Epub 2006 Mar 28. Cardiovasc Res. 2006. PMID: 16650839 Review.
-
Zebrin II expressing Purkinje cell phenotype-related and -unrelated cerebellar abnormalities in Cav2.1 mutant, rolling mouse Nagoya.ScientificWorldJournal. 2010 Oct 12;10:2032-8. doi: 10.1100/tsw.2010.205. ScientificWorldJournal. 2010. PMID: 20953553 Free PMC article. Review.
Cited by
-
Role of Ca2+ in healthy and pathologic cardiac function: from normal excitation-contraction coupling to mutations that cause inherited arrhythmia.Arch Toxicol. 2023 Jan;97(1):73-92. doi: 10.1007/s00204-022-03385-0. Epub 2022 Oct 10. Arch Toxicol. 2023. PMID: 36214829 Free PMC article. Review.
-
BDNF-TrkB signaling in oxytocin neurons contributes to maternal behavior.Elife. 2018 Sep 7;7:e33676. doi: 10.7554/eLife.33676. Elife. 2018. PMID: 30192229 Free PMC article.
-
A Mutation in the ANK2 Gene Causing ASD and a Review of the Literature.Mol Genet Genomic Med. 2025 Mar;13(3):e70083. doi: 10.1002/mgg3.70083. Mol Genet Genomic Med. 2025. PMID: 40035441 Free PMC article. Review.
-
Mechanisms underlying the role of ankyrin-B in cardiac and neurological health and disease.Front Cardiovasc Med. 2022 Aug 4;9:964675. doi: 10.3389/fcvm.2022.964675. eCollection 2022. Front Cardiovasc Med. 2022. PMID: 35990955 Free PMC article. Review.
-
Physical and functional convergence of the autism risk genes Scn2a and Ank2 in neocortical pyramidal cell dendrites.Neuron. 2024 Apr 3;112(7):1133-1149.e6. doi: 10.1016/j.neuron.2024.01.003. Epub 2024 Jan 29. Neuron. 2024. PMID: 38290518 Free PMC article.
References
-
- Cheever T. R., Ervasti J. M. (2013) Actin isoforms in neuronal development and function. Int. Rev. Cell Mol. Biol. 301, 157–213 - PubMed
-
- Bennett V., Healy J. (2008) Organizing the fluid membrane bilayer: diseases linked to spectrin and ankyrin. Trends Mol. Med. 14, 28–36 - PubMed
-
- Dunlap K., Luebke J. I., Turner T. J. (1995) Exocytotic Ca2+ channels in mammalian central neurons. Trends Neurosci. 18, 89–98 - PubMed
-
- Catterall W. A. (2000) Structure and regulation of voltage-gated Ca2+ channels. Annu. Rev. Cell Dev. Biol. 16, 521–555 - PubMed
-
- Clapham D. E. (1995) Calcium signaling. Cell 80, 259–268 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
