

A TDG/CBP/RARα ternary complex mediates the retinoic acid-dependent expression of DNA methylation-sensitive genes
- PMID: 24394593
- PMCID: PMC4411359
- DOI: 10.1016/j.gpb.2013.11.001
A TDG/CBP/RARα ternary complex mediates the retinoic acid-dependent expression of DNA methylation-sensitive genes
Abstract
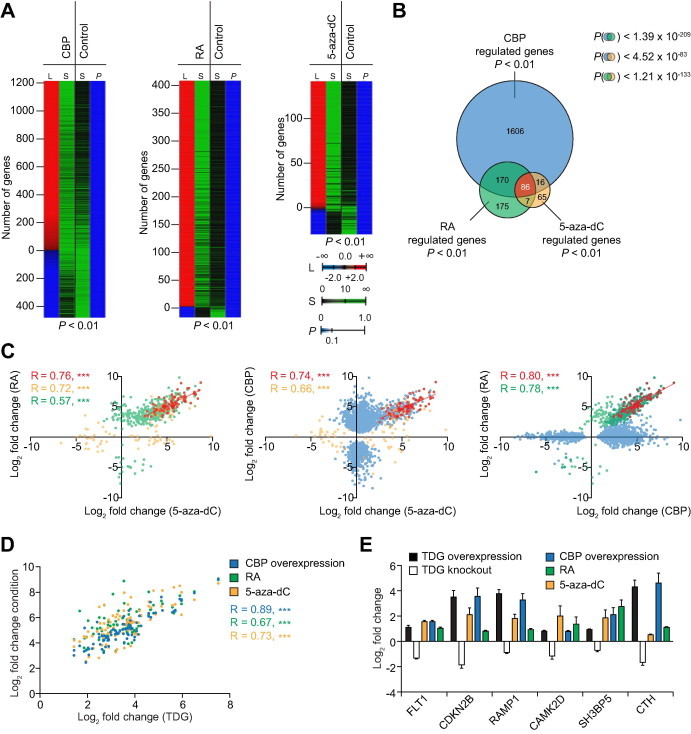
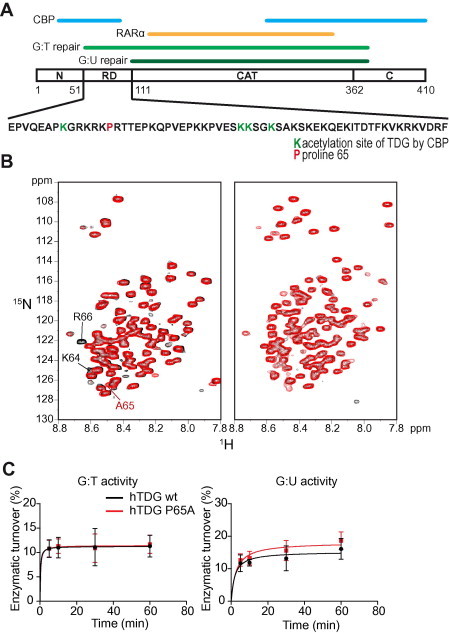
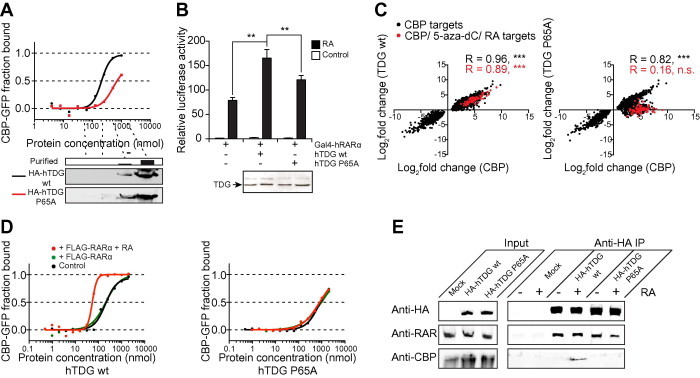
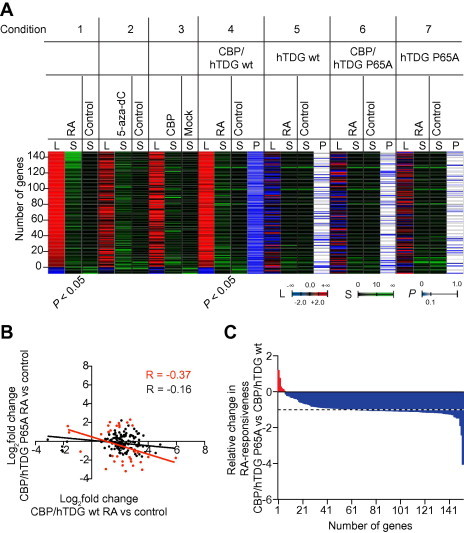
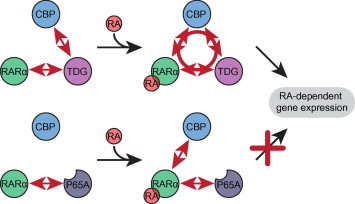
The thymine DNA glycosylase (TDG) is a multifunctional enzyme, which is essential for embryonic development. It mediates the base excision repair (BER) of G:T and G:U DNA mismatches arising from the deamination of 5-methyl cytosine (5-MeC) and cytosine, respectively. Recent studies have pointed at a role of TDG during the active demethylation of 5-MeC within CpG islands. TDG interacts with the histone acetylase CREB-binding protein (CBP) to activate CBP-dependent transcription. In addition, TDG also interacts with the retinoic acid receptor α (RARα), resulting in the activation of RARα target genes. Here we provide evidence for the existence of a functional ternary complex containing TDG, CBP and activated RARα. Using global transcriptome profiling, we uncover a coupling of de novo methylation-sensitive and RA-dependent transcription, which coincides with a significant subset of CBP target genes. The introduction of a point mutation in TDG, which neither affects overall protein structure nor BER activity, leads to a significant loss in ternary complex stability, resulting in the deregulation of RA targets involved in cellular networks associated with DNA replication, recombination and repair. We thus demonstrate for the first time a direct coupling of TDG's epigenomic and transcription regulatory function through ternary complexes with CBP and RARα.
Keywords: CREB-binding protein; Cytosine DNA methylation; Epigenomics; Retinoic acid receptor α; Thymine DNA glycosylase; Transcription regulation.
Copyright © 2014. Production and hosting by Elsevier Ltd.
Figures

Similar articles
-
The thymine-DNA glycosylase regulatory domain: residual structure and DNA binding.Biochemistry. 2008 Jun 24;47(25):6519-30. doi: 10.1021/bi7022283. Biochemistry. 2008. PMID: 18512959
-
Thymine DNA glycosylase is essential for active DNA demethylation by linked deamination-base excision repair.Cell. 2011 Jul 8;146(1):67-79. doi: 10.1016/j.cell.2011.06.020. Epub 2011 Jun 30. Cell. 2011. PMID: 21722948 Free PMC article.
-
Regulation of Active DNA Demethylation through RAR-Mediated Recruitment of a TET/TDG Complex.Cell Rep. 2017 May 23;19(8):1685-1697. doi: 10.1016/j.celrep.2017.05.007. Cell Rep. 2017. PMID: 28538185
-
Role of base excision repair in maintaining the genetic and epigenetic integrity of CpG sites.DNA Repair (Amst). 2015 Aug;32:33-42. doi: 10.1016/j.dnarep.2015.04.011. Epub 2015 May 1. DNA Repair (Amst). 2015. PMID: 26021671 Free PMC article. Review.
-
DNA demethylation by TDG.Epigenomics. 2012 Aug;4(4):459-67. doi: 10.2217/epi.12.36. Epigenomics. 2012. PMID: 22920184 Free PMC article. Review.
Cited by
-
The Role of Thymine DNA Glycosylase in Transcription, Active DNA Demethylation, and Cancer.Cancers (Basel). 2022 Feb 1;14(3):765. doi: 10.3390/cancers14030765. Cancers (Basel). 2022. PMID: 35159032 Free PMC article. Review.
-
Multifaceted roles for thymine DNA glycosylase in embryonic development and human carcinogenesis.Acta Biochim Biophys Sin (Shanghai). 2016 Jan;48(1):82-9. doi: 10.1093/abbs/gmv083. Epub 2015 Sep 14. Acta Biochim Biophys Sin (Shanghai). 2016. PMID: 26370152 Free PMC article. Review.
-
Linking functions: an additional role for an intrinsically disordered linker domain in the transcriptional coactivator CBP.Sci Rep. 2017 Jul 5;7(1):4676. doi: 10.1038/s41598-017-04611-x. Sci Rep. 2017. PMID: 28680062 Free PMC article.
-
Interaction with the DNA Repair Protein Thymine DNA Glycosylase Regulates Histone Acetylation by p300.Biochemistry. 2016 Dec 13;55(49):6766-6775. doi: 10.1021/acs.biochem.6b00841. Epub 2016 Dec 1. Biochemistry. 2016. PMID: 27951654 Free PMC article.
-
The current state of eukaryotic DNA base damage and repair.Nucleic Acids Res. 2015 Dec 2;43(21):10083-101. doi: 10.1093/nar/gkv1136. Epub 2015 Oct 30. Nucleic Acids Res. 2015. PMID: 26519467 Free PMC article. Review.
References
-
- Schär P., Fritsch O. DNA repair and the control of DNA methylation. Prog Drug Res. 2011;67:51–68. - PubMed
-
- Ehrlich M., Zhang X.Y., Inamdar N.M. Spontaneous deamination of cytosine and 5-methylcytosine residues in DNA and replacement of 5-methylcytosine residues with cytosine residues. Mutat Res. 1990;238:277–286. - PubMed
-
- Krokan H.E., Drabløs F., Slupphaug G. Uracil in DNA – occurrence, consequences and repair. Oncogene. 2001;21:8935–8948. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
