

A molecular brake controls the magnitude of long-term potentiation
- PMID: 24394804
- PMCID: PMC3895372
- DOI: 10.1038/ncomms4051
A molecular brake controls the magnitude of long-term potentiation
Abstract
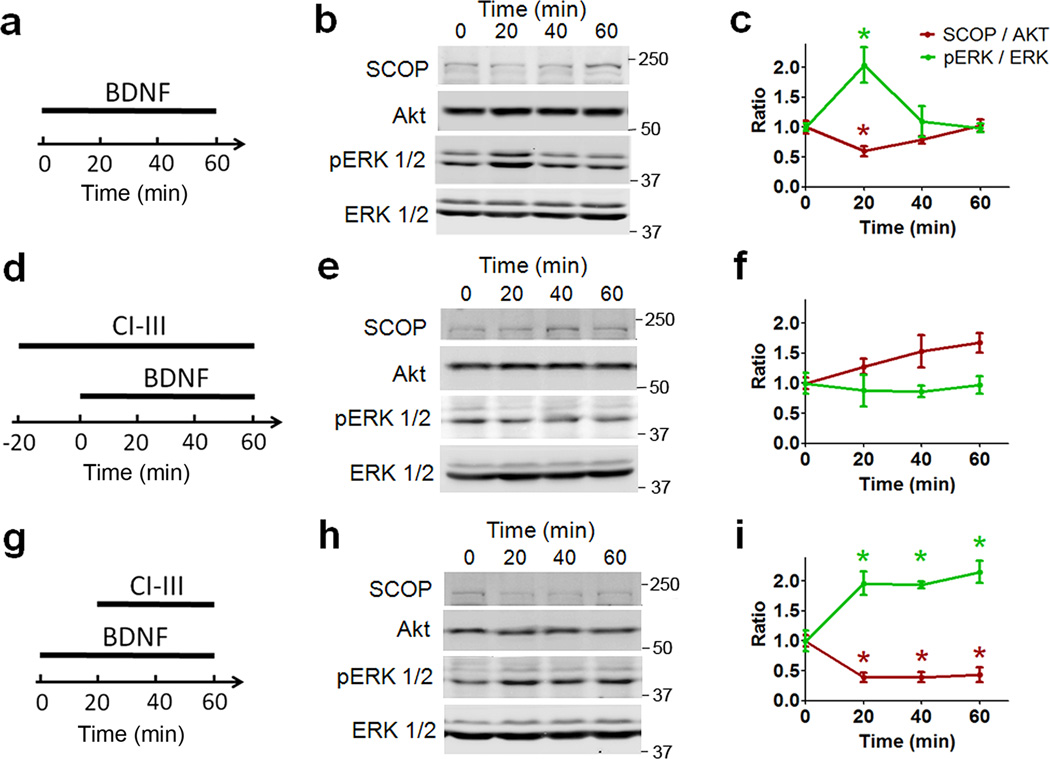
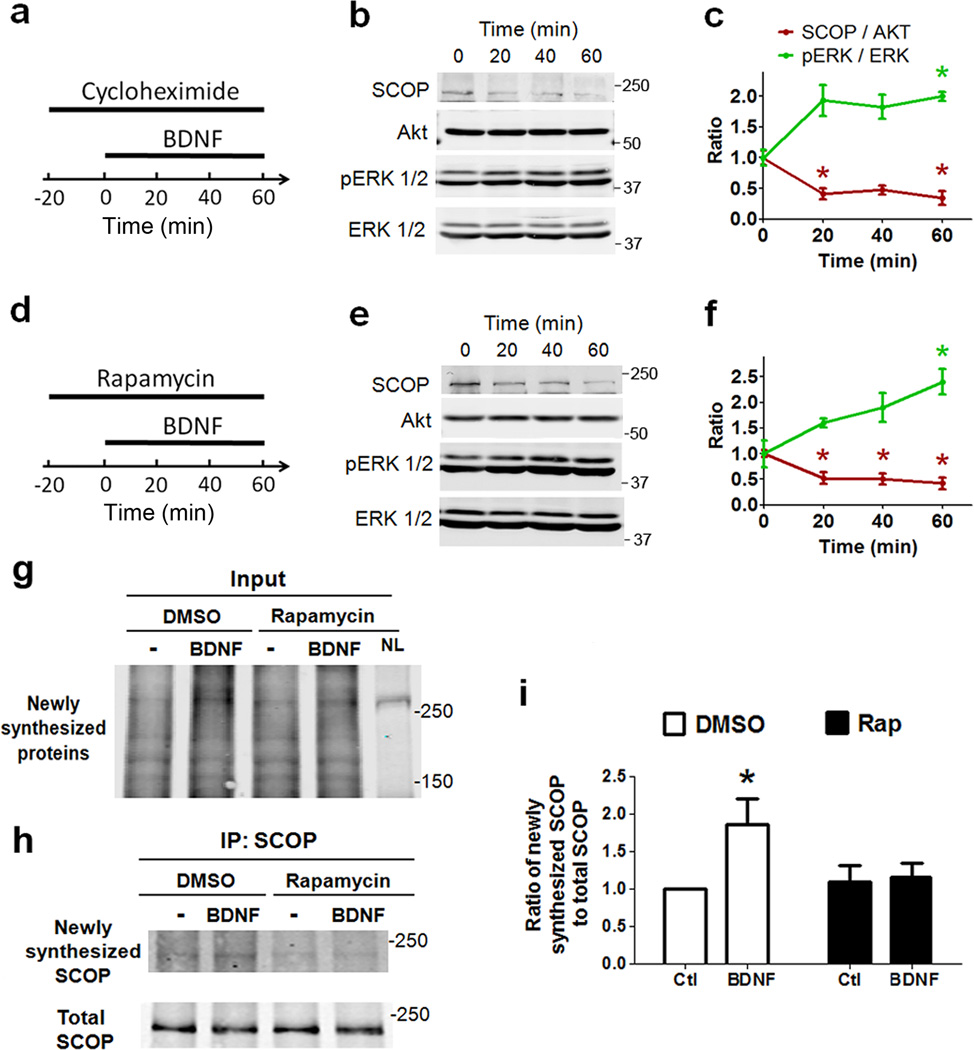
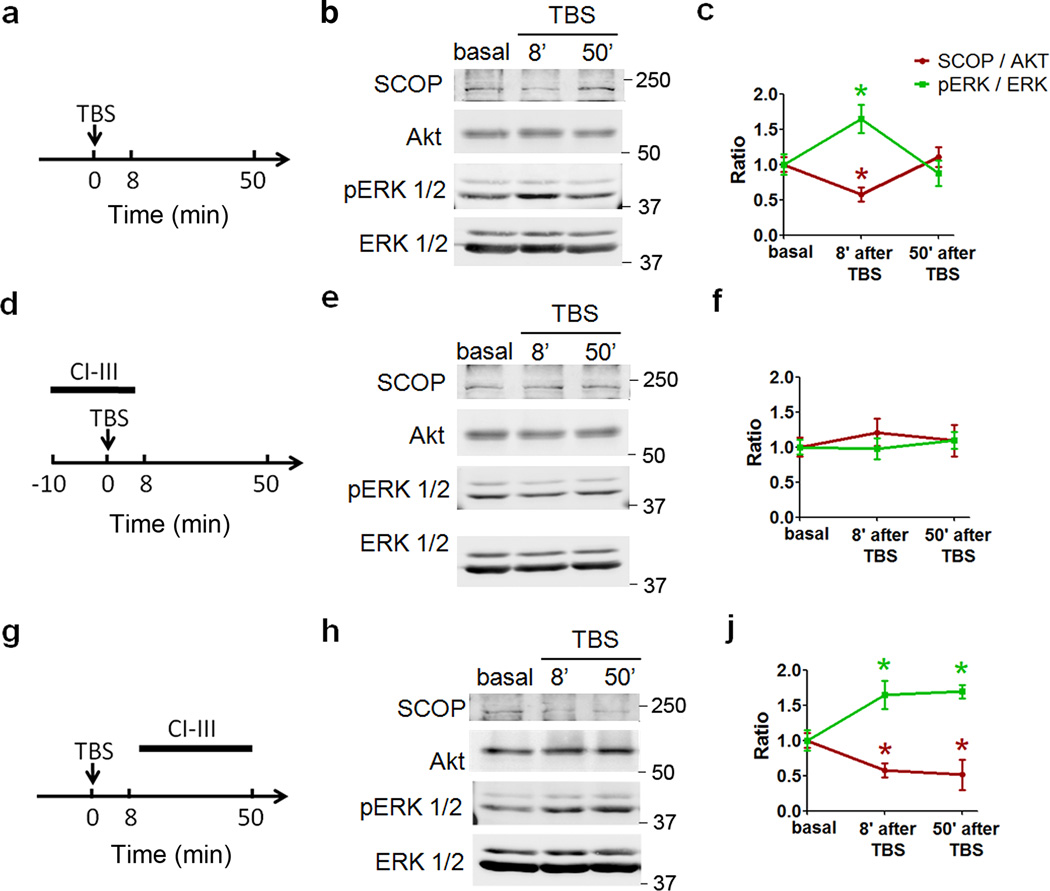
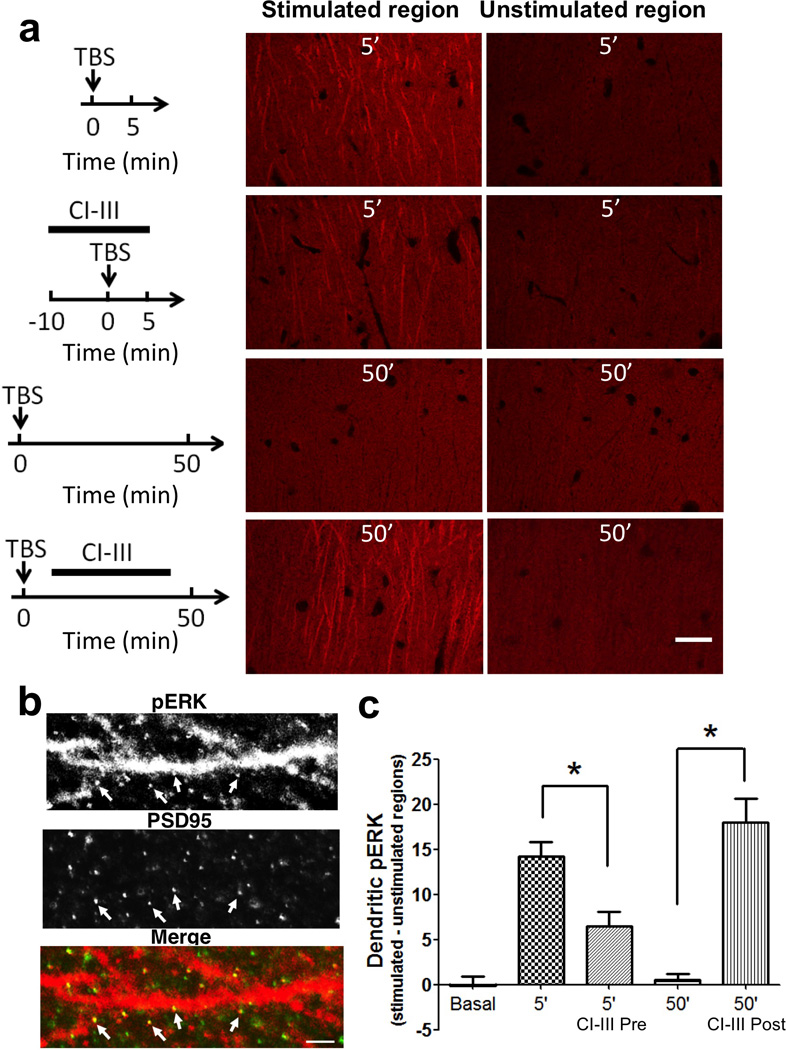
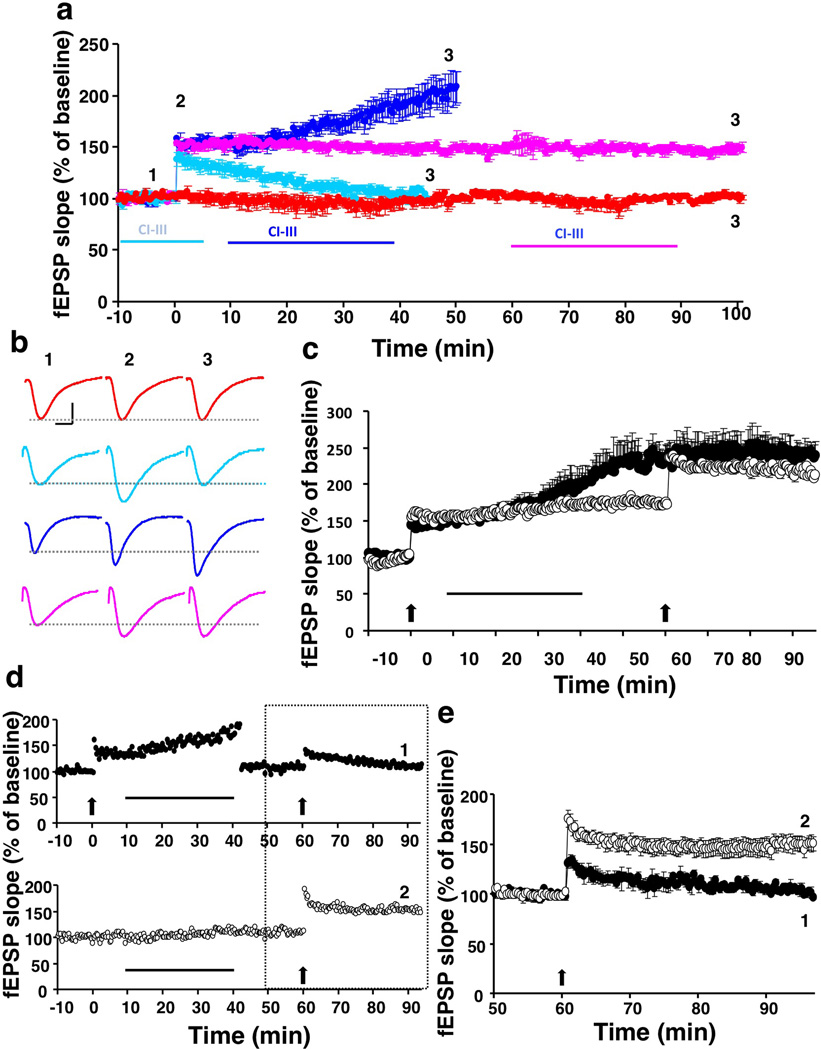
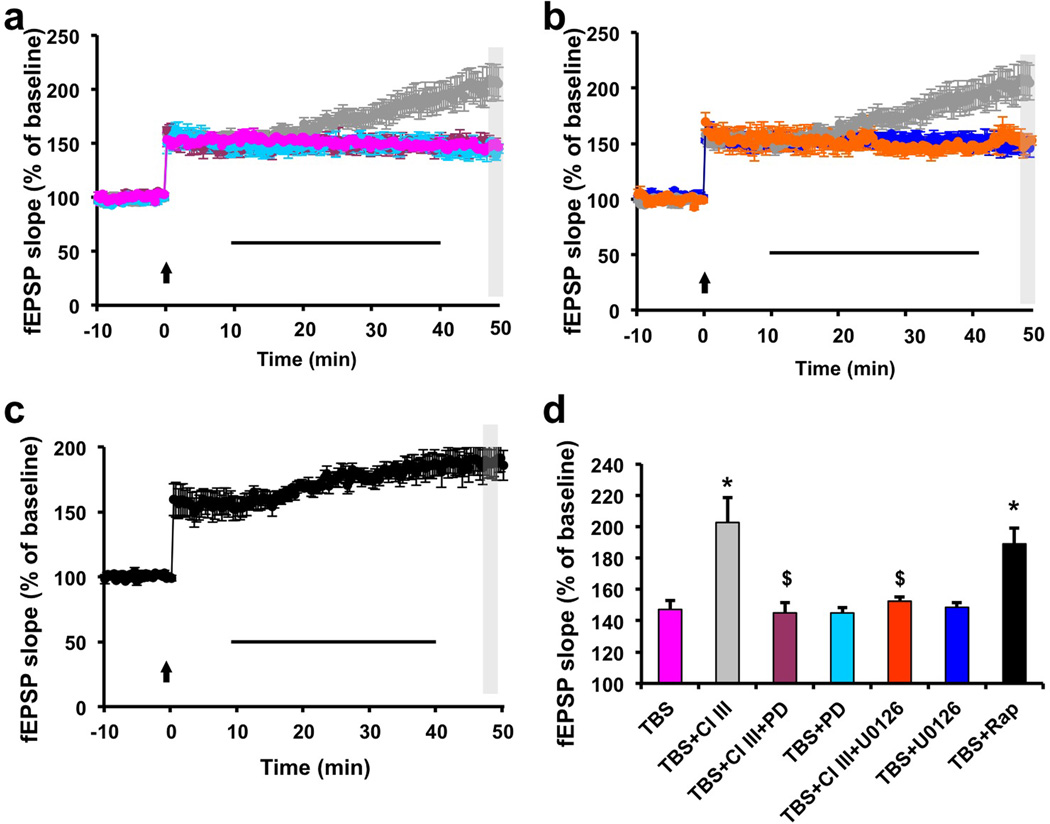
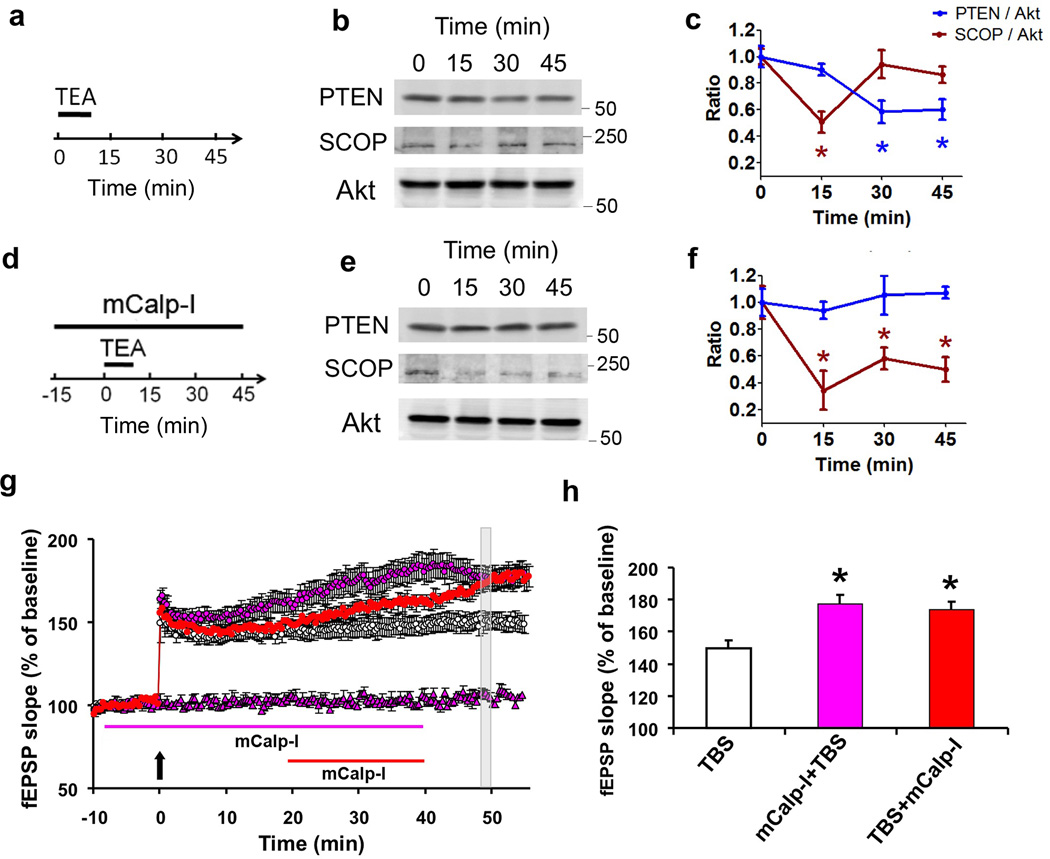
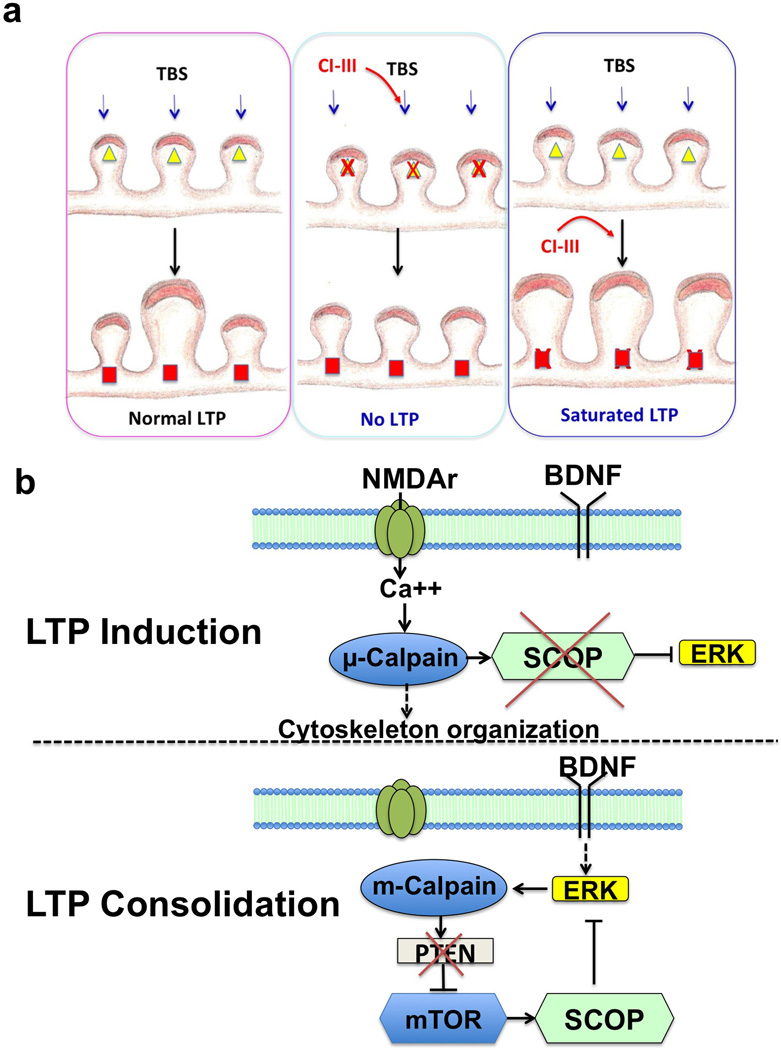
Overexpression of suprachiasmatic nucleus circadian oscillatory protein (SCOP), a negative ERK regulator, blocks long-term memory encoding. Inhibition of calpain-mediated SCOP degradation also prevents the formation of long-term memory, suggesting rapid SCOP breakdown is necessary for memory encoding. However, whether SCOP levels also control the magnitude of long-term synaptic plasticity is unknown. Here we show that following synaptic activity-induced SCOP degradation, SCOP is rapidly replaced via mTOR-mediated protein synthesis. We further show that early SCOP degradation is specifically catalysed by μ-calpain, whereas late SCOP resynthesis is mediated by m-calpain. We propose that μ-calpain promotes long-term potentiation induction by degrading SCOP and activating ERK, whereas m-calpain activation limits the magnitude of potentiation by terminating the ERK response via enhanced SCOP synthesis. This unique braking mechanism could account for the advantages of spaced versus massed training in the formation of long-term memory.
Figures
References
-
- Kelleher RJ, 3rd, Govindarajan A, Jung HY, Kang H, Tonegawa S. Translational control by MAPK signaling in long-term synaptic plasticity and memory. Cell. 2004;116:467–479. - PubMed
-
- Winder DG, et al. ERK plays a regulatory role in induction of LTP by theta frequency stimulation and its modulation by beta-adrenergic receptors. Neuron. 1999;24:715–726. - PubMed
-
- Shimizu K, Okada M, Nagai K, Fukada Y. Suprachiasmatic nucleus circadian oscillatory protein, a novel binding partner of K-Ras in the membrane rafts, negatively regulates MAPK pathway. J Biol Chem. 2003;278:14920–14925. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
