

The multifork Escherichia coli chromosome is a self-duplicating and self-segregating thermodynamic ring polymer
- PMID: 24395248
- PMCID: PMC3894414
- DOI: 10.1101/gad.231050.113
The multifork Escherichia coli chromosome is a self-duplicating and self-segregating thermodynamic ring polymer
Abstract
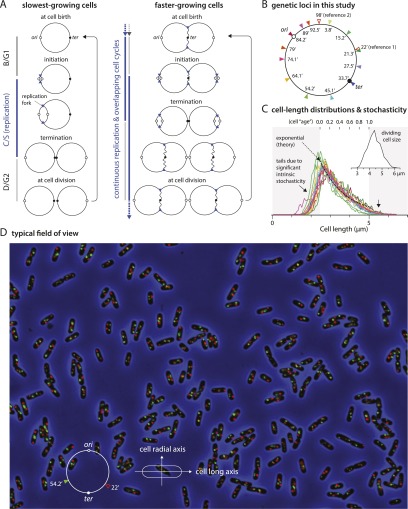
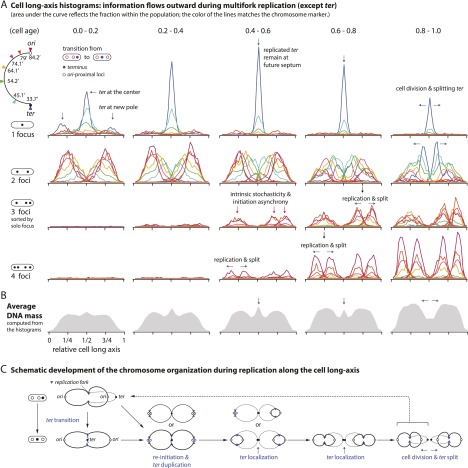
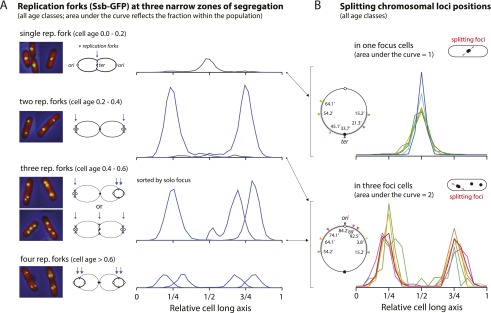
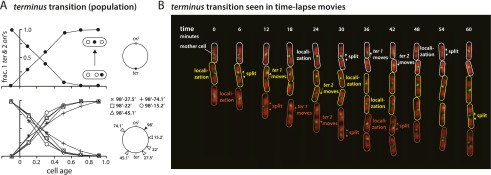
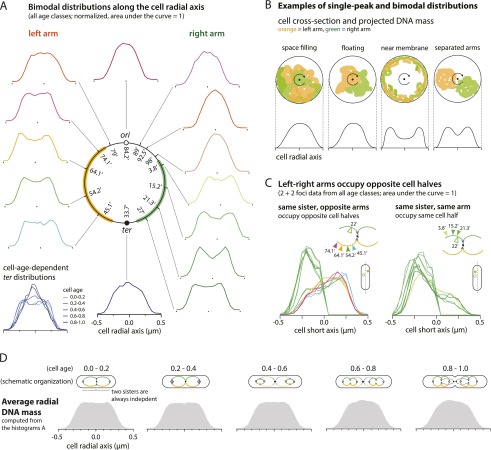
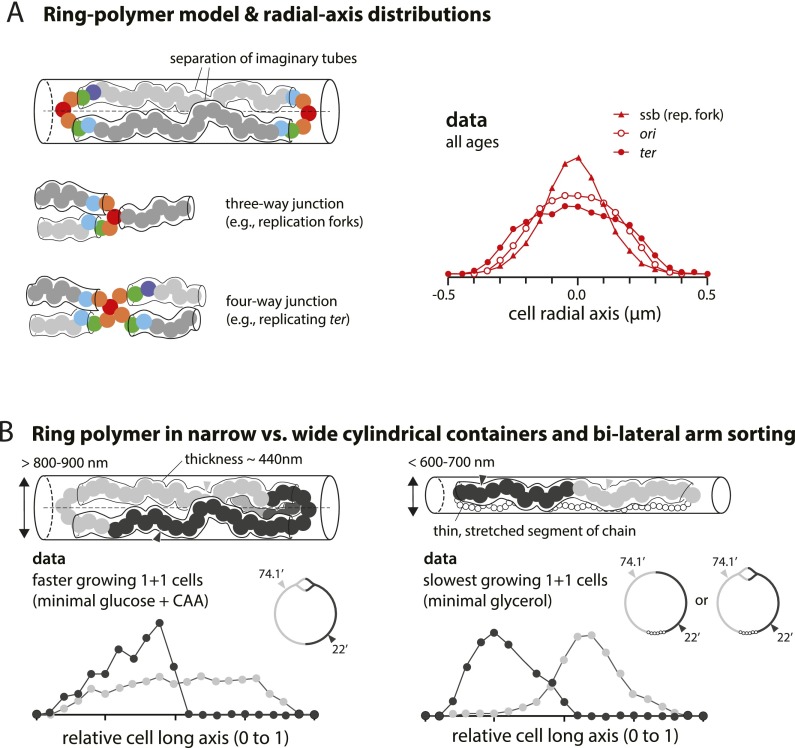
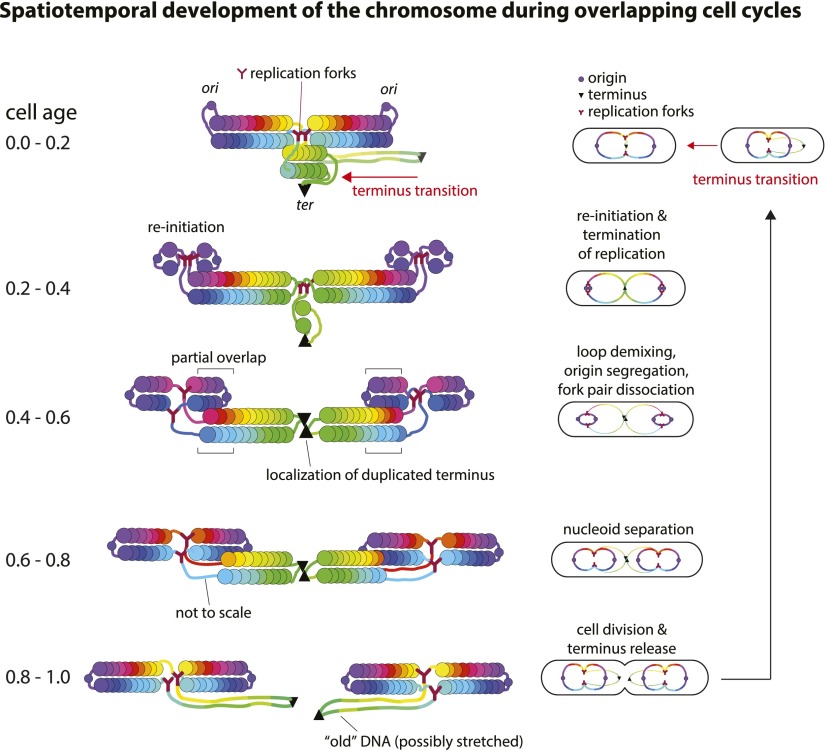
At all but the slowest growth rates, Escherichia coli cell cycles overlap, and its nucleoid is segregated to daughter cells as a forked DNA circle with replication ongoing-a state fundamentally different from eukaryotes. We have solved the chromosome organization, structural dynamics, and segregation of this constantly replicating chromosome. It is locally condensed to form a branched donut, compressed so that the least replicated DNA spans the cell center and the newest DNA extends toward the cell poles. Three narrow zones at the cell center and quarters contain both the replication forks and nascent DNA and serve to segregate the duplicated chromosomal information as it flows outward. The overall pattern is smoothly self-replicating, except when the duplicated terminus region is released from the septum and recoils to the center of a sister nucleoid. In circular cross-section of the cell, the left and right arms of the chromosome form separate, parallel structures that lie in each cell half along the radial cell axis. In contrast, replication forks and origin and terminus regions are found mostly at the center of the cross section, balanced by the parallel chromosome arms. The structure is consistent with the model in which the nucleoid is a constrained ring polymer that develops by spontaneous thermodynamics. The ring polymer pattern extrapolates to higher growth rates and also provides a structural basis for the form of the chromosome during very slow growth.
Keywords: bacterial cell cycle; bacterial chromosome; chromosome organization; chromosome segregation; multifork replication.
Figures
References
-
- Adachi S, Kohiyama M, Onogi T, Hiraga S 2005. Localization of replication forks in wild-type and mukB mutant cells of Escherichia coli. Mol Genet Genomics 274: 264–271 - PubMed
-
- Adachi S, Fukushima T, Hiraga S 2008. Dynamic events of sister chromosomes in the cell cycle of Escherichia coli. Genes Cells 13: 181–197 - PubMed
-
- Bigot S, Sivanathan V, Possoz C, Barre FX, Cornet F 2007. FtsK, a literate chromosome segregation machine. Mol Microbiol 64: 1434–1441 - PubMed