

Prenatal cerebral ischemia triggers dysmaturation of caudate projection neurons
- PMID: 24395459
- PMCID: PMC4013245
- DOI: 10.1002/ana.24100
Prenatal cerebral ischemia triggers dysmaturation of caudate projection neurons
Abstract
Objective: Recently, we reported that the neocortex displays impaired growth after transient cerebral hypoxia-ischemia (HI) at preterm gestation that is unrelated to neuronal death but is associated with decreased dendritic arbor complexity of cortical projection neurons. We hypothesized that these morphological changes constituted part of a more widespread neuronal dysmaturation response to HI in the caudate nucleus (CN), which contributes to motor and cognitive disability in preterm survivors.
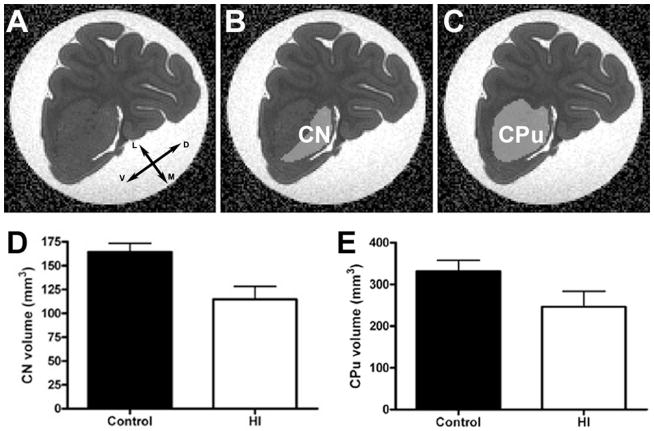
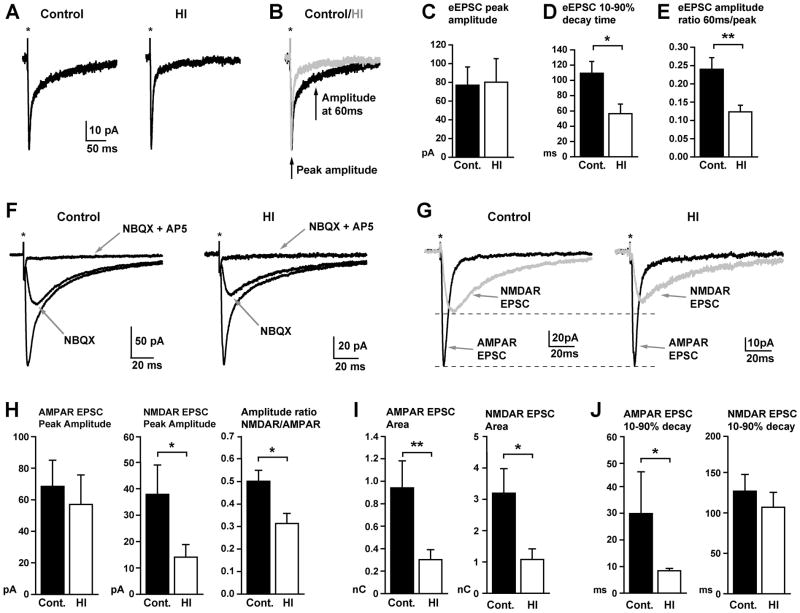
Methods: Ex vivo magnetic resonance imaging (MRI), immunohistochemistry, and Golgi staining defined CN growth, cell death, proliferation, and dendritic maturation in preterm fetal sheep 4 weeks after HI. Patch-clamp recording was used to analyze glutamatergic synaptic currents in CN neurons.
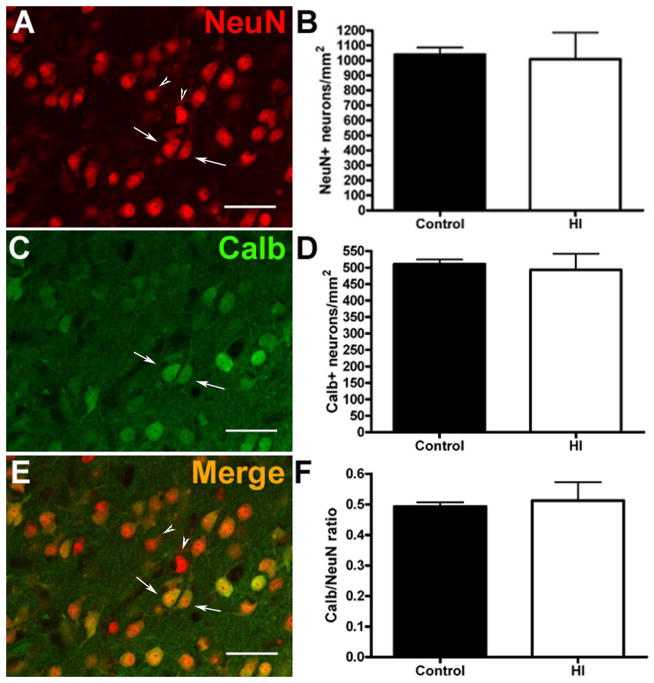
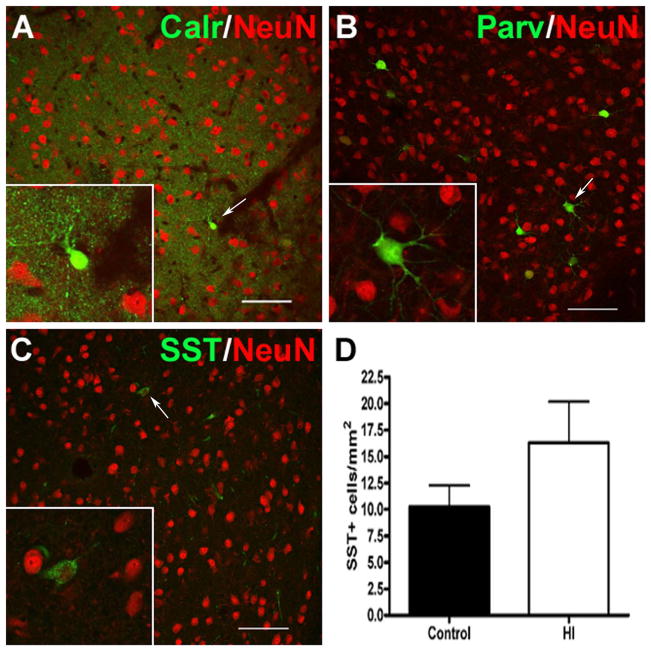
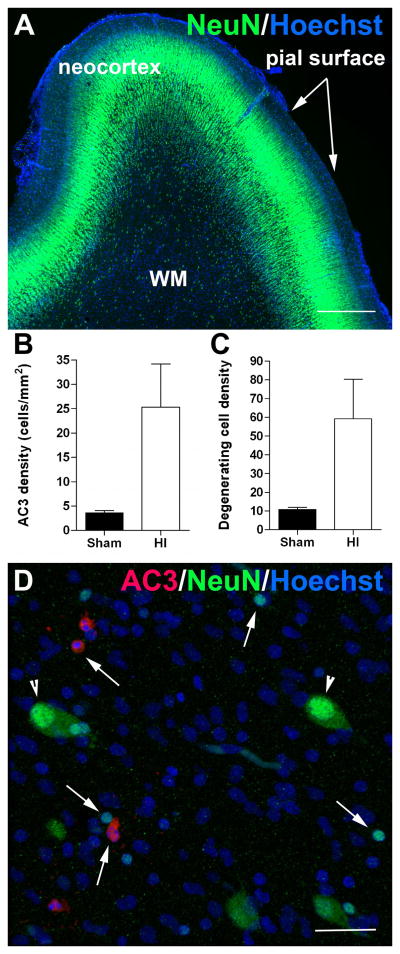
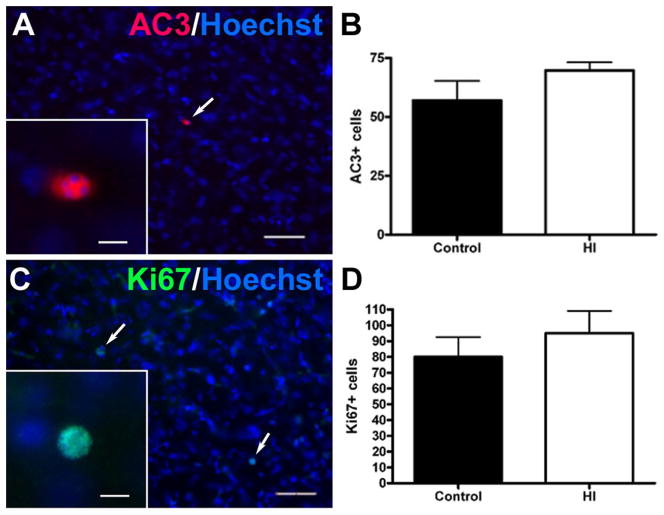
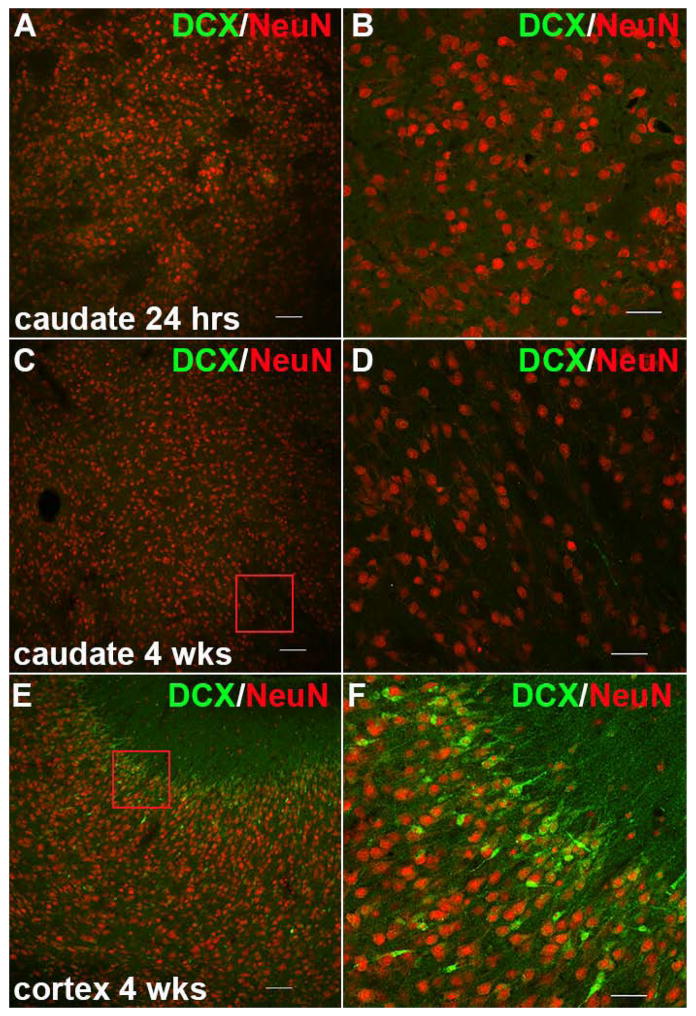
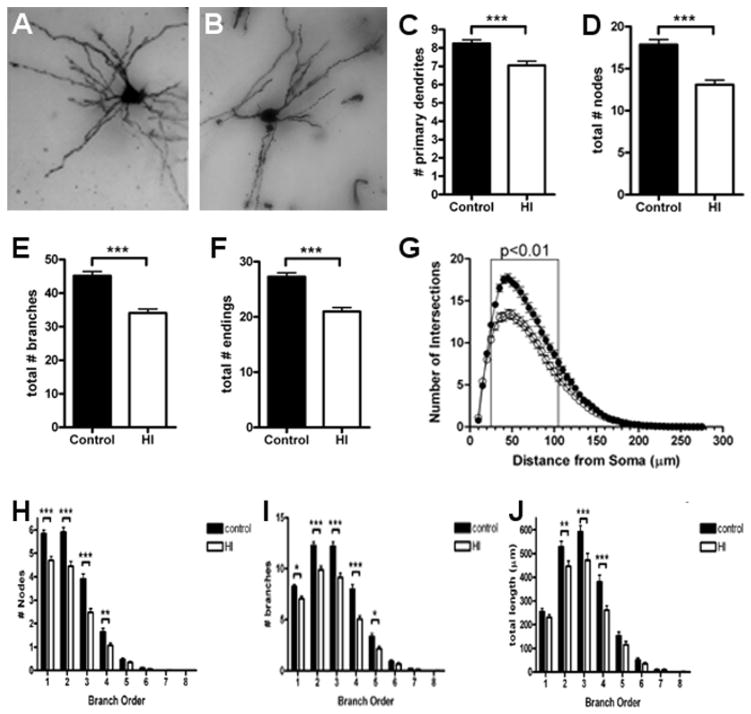
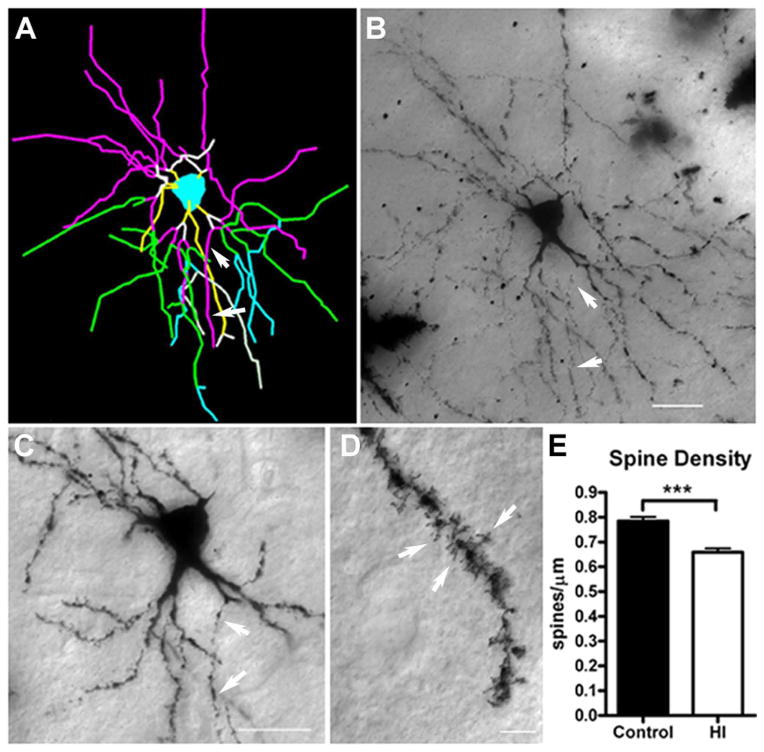
Results: MRI-defined growth of the CN was reduced after ischemia compared to controls. However, no significant acute or delayed neuronal death was seen in the CN or white matter. Nor was there significant loss of calbindin-positive medium spiny projection neurons (MSNs) or CN interneurons expressing somatostatin, calretinin, parvalbumin, or tyrosine hydroxylase. Morphologically, ischemic MSNs showed a markedly immature dendritic arbor, with fewer dendritic branches, nodes, endings, and spines. The magnitude and kinetics of synaptic currents, and the relative contribution of glutamate receptor subtypes in the CN were significantly altered.
Interpretation: The marked MSN dendritic and functional abnormalities after preterm cerebral HI, despite the marked resistance of immature CN neurons to cell death, are consistent with widespread susceptibility of projection neurons to HI-induced dysmaturation. These global disturbances in dendritic maturation and glutamatergic synaptic transmission suggest a new mechanism for long-term motor and behavioral disabilities in preterm survivors via widespread disruption of neuronal connectivity.
© 2014 American Neurological Association.
Conflict of interest statement
Nothing to report.
Figures
References
-
- Allen M. Outcomes after brain injury in the preterm infant. In: Miller S, Shevell M, editors. Acquired Brain Injury in the Fetus and Newborn. London: Mac Keith Press; 2012. pp. 99–120.
-
- Keunen K, Kersbergen KJ, Groenendaal F, Isgum I, de Vries LS, Benders MJ. Brain tissue volumes in preterm infants: prematurity, perinatal risk factors and neurodevelopmental outcome: a systematic review. J Matern Fetal Neonatal Med. 2012;25 (Suppl 1):89–100. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
