

Review
doi: 10.1083/jcb.201310021.
Adaptor proteins involved in polarized sorting
Affiliations
- PMID: 24395635
- PMCID: PMC3882786
- DOI: 10.1083/jcb.201310021
Item in Clipboard
Review
Adaptor proteins involved in polarized sorting
J Cell Biol.
.
Abstract
Polarized cells such as epithelial cells and neurons exhibit different plasma membrane domains with distinct protein compositions. Recent studies have shown that sorting of transmembrane proteins to the basolateral domain of epithelial cells and the somatodendritic domain of neurons is mediated by recognition of signals in the cytosolic domains of the proteins by adaptors. These adaptors are components of protein coats associated with the trans-Golgi network and/or recycling endosomes. The clathrin-associated adaptor protein 1 (AP-1) complex plays a preeminent role in this process, although other adaptors and coat proteins, such as AP-4, ARH, Numb, exomer, and retromer, have also been implicated.
Figures
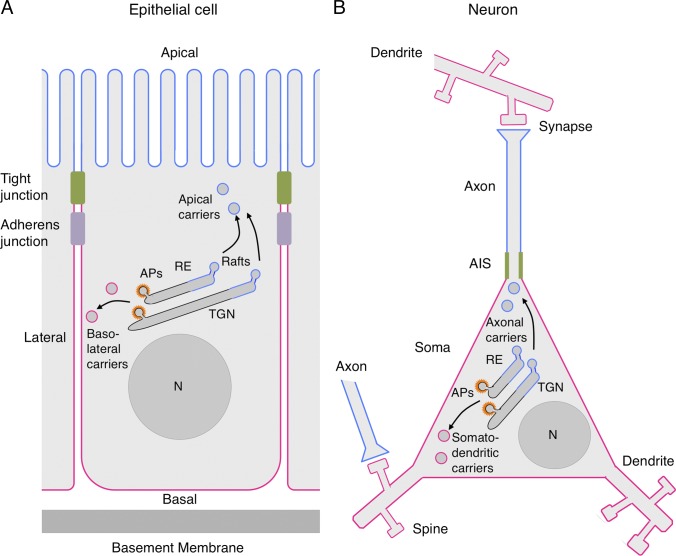
Schematic representation of a polarized epithelial cell and a neuron. (A) The scheme depicts a polarized epithelial cell flanked by two neighboring cells. Tight junctions block the passage of substances through the intercellular space and also serve as a boundary between the apical (blue) and basolateral (red) domains of the plasma membrane. Adherens junctions hold cells together and regulate their proliferation. Vesicular carriers that transport proteins to the apical and basolateral plasma membrane domains are shown to emanate from both the trans-Golgi network (TGN) and recycling endosomes (RE), reflecting polarized sorting in both the biosynthetic and recycling pathways, respectively. Apical carriers are shown to arise from cholesterol- and glycosphingolipid-enriched “raft” domains, whereas basolateral carriers arise from adaptor protein (AP)–enriched “coat” domains. (B) Depiction of a neuron making synapses with two other neurons. Synapses are shown as contacts between an axon terminal and a dendritic spine, but post-synaptic sites also occur on the dendritic shaft or soma. The boundary between the axonal (blue) and somatodendritic (red) domains of the plasma membrane is at the axon initial segment (AIS, green). Axonal and somatodendritic carriers are shown emanating from the TGN and RE, as in epithelial cells. Although both schemes depict polarized sorting occurring by segregation into two types of transport carriers at the TGN/RE, selective retention at or removal from a particular domain of the plasma membrane, as well as transcytosis, also determine polarized distribution of some proteins in both epithelial cells and neurons (Garrido et al., 2001; Gan et al., 2002; Sampo et al., 2003
Anderson et al., 2005; Yap et al., 2008). A trans-endosomal route involving transport from the TGN to RE in the biosynthetic pathway has also been demonstrated (Ang et al., 2004). N, nucleus.

Signals and adaptor proteins involved in polarized sorting. (A) Sequences of the cytosolic tails of the indicated proteins, with critical residues of polarized sorting signals highlighted in red. Signals are categorized as YXXØ, [DE]XXXL[LI], or noncanonical. The transmembrane domain (TM) and the number of additional residues in each tail are indicated. For the TfR, the basolateral sorting signal is GDNS, but the somatodendritic sorting signal is YTRF. The LDLR has a proximal signal with one tyrosine residue and an acidic cluster (EDE) and a distal signal with two tyrosine residues and another acidic cluster (EED). (B) Adaptor proteins that have been implicated in polarized sorting. AP-1 and AP-4 are composed of four homologous subunits, as reflected by the color scheme. The core, hinge, and ear domains of the AP complexes and the PTB domain of ARH and Numb are indicated. Folded regions are represented by geometric shapes and disordered regions by wavy lines. The AP-2, AP-3, AP-5, and COPI-F complexes are homologous to AP-1 and AP-4, but to date they have not been directly implicated in polarized sorting. Three of the AP-1 subunits occur as isoforms encoded by different genes: γ1 and γ2; μ1A and μ1B; and σ1A, σ1B, and σ1C. PM, plasma membrane. (C) Schematic representation of a clathrin-coated bud on the TGN/RE containing AP-1, Numb, and ARH as adaptor proteins. Arf proteins recruit AP-1 to membranes and promote its conformational activation. Clathrin triskelia polymerize onto the adaptor proteins to form a polyhedral coat. Cargos are gathered into the coated buds by interaction of sorting signals with the adaptor proteins.
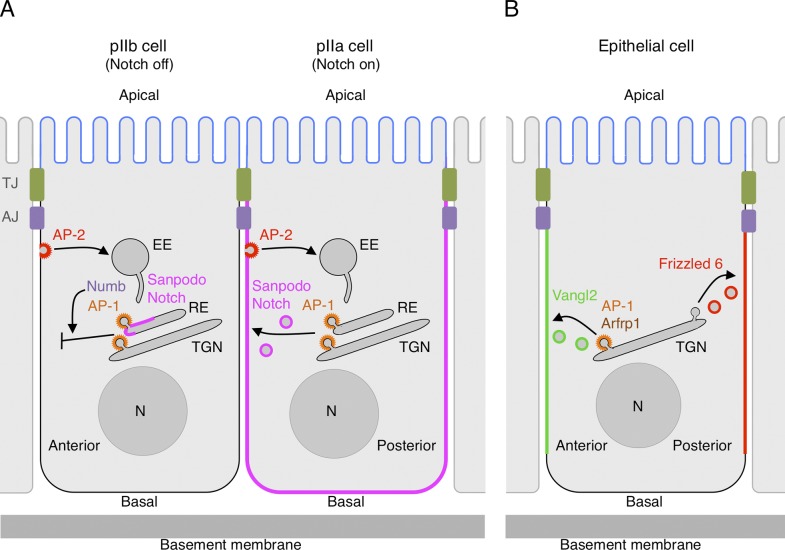
Role of AP-1 in cell fate specification and planar cell polarity. (A) Asymmetric inheritance of Numb in Drosophila sensory organ precursor (SOP) daughter cells determines AP-1–dependent inhibition of Sanpodo and Notch recycling in the pIIb cell (Numb positive) and allows AP-1–dependent recycling of Sanpodo and Notch in the pIIa cell (Numb negative; Benhra et al., 2011; Cotton et al., 2013). Notch signaling is thus inhibited in the pIIb cell and activated in the pIIa cell. The related AP-2 complex mediates Notch and Sanpodo internalization, but Numb does not appear to regulate this process (Cotton et al., 2013; Couturier et al., 2013). The anterior or posterior orientation of the SOP daughter cells in the plane of the notum is indicated. EE, early endosome. (B) AP-1 in association with the Arf-like protein Arfrp1 mediates export of the planar cell polarity (PCP) regulator Vangl2 from the TGN to the anterior (or proximal) surface of the lateral membrane domain in epithelial cells (Y. Guo et al., 2013). Another PCP regulator, Frizzled 6, is targeted to the posterior (or distal) surface by an unknown mechanism. AJ, adherens junction; EE, early endosome; N, nucleus; TJ, tight junction.
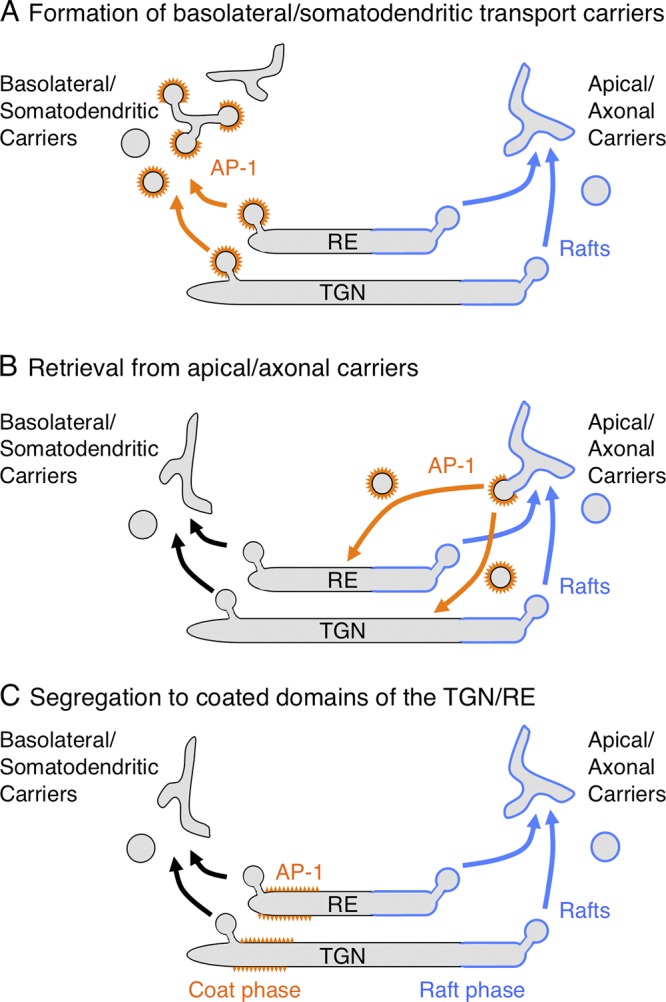
Models of AP-1 function in polarized sorting at the TGN and RE. The schemes depict three models for the function of AP-1 in sorting to the basolateral domain of epithelial cells and the somatodendritic domain of neurons. In model A, AP-1 sorts cargos into clathrin-coated vesicles or pleiomorphic transport carriers at the TGN/RE. These intermediates lose their coats and eventually deliver their cargos to the basolateral or somatodendritic plasma membrane domains. In model B, AP-1 removes basolateral or somatodendritic cargos from apical or axonal transport carriers to return them to the TGN/RE. In model C, AP-1 segregates basolateral or somatodendritic cargos into a “coat phase” of the TGN/RE, away from the “raft phase” that gives rise to apical or axonal carriers.
References
-
- Abou Jamra R., Philippe O., Raas-Rothschild A., Eck S.H., Graf E., Buchert R., Borck G., Ekici A., Brockschmidt F.F., Nöthen M.M., et al. 2011. Adaptor protein complex 4 deficiency causes severe autosomal-recessive intellectual disability, progressive spastic paraplegia, shy character, and short stature. Am. J. Hum. Genet. 88:788–795 10.1016/j.ajhg.2011.04.019 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
