

New insights into lineage restriction of mammary gland epithelium using parity-identified mammary epithelial cells
- PMID: 24398145
- PMCID: PMC3978646
- DOI: 10.1186/bcr3593
New insights into lineage restriction of mammary gland epithelium using parity-identified mammary epithelial cells
Abstract
Introduction: Parity-identified mammary epithelial cells (PI-MECs) are an interesting cellular subset because they survive involution and are a presumptive target for transformation by human epidermal growth factor receptor 2 (HER2)/neu in mammary tumors. Depending on the type of assay, PI-MECs have been designated lobule-restricted progenitors or multipotent stem/progenitor cells. PI-MECs were reported to be part of the basal population of mammary epithelium based on flow cytometry. We investigated the cellular identity and lineage potential of PI-MECs in intact mammary glands.
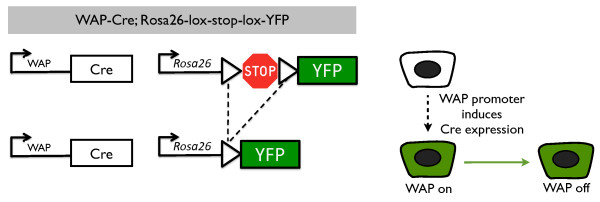
Methods: We performed a quantitative and qualitative analysis of the contribution of PI-MECs to mammary epithelial cell lineages in pregnant and involuted mammary glands by immunohistochemistry, fluorescence-activated cells sorting (FACS), and quantitative polymerase chain reaction. PI-MECs were labeled by the activation of Whey Acidic Protein (WAP)-Cre during pregnancy that results in permanent expression of yellow fluorescent protein.
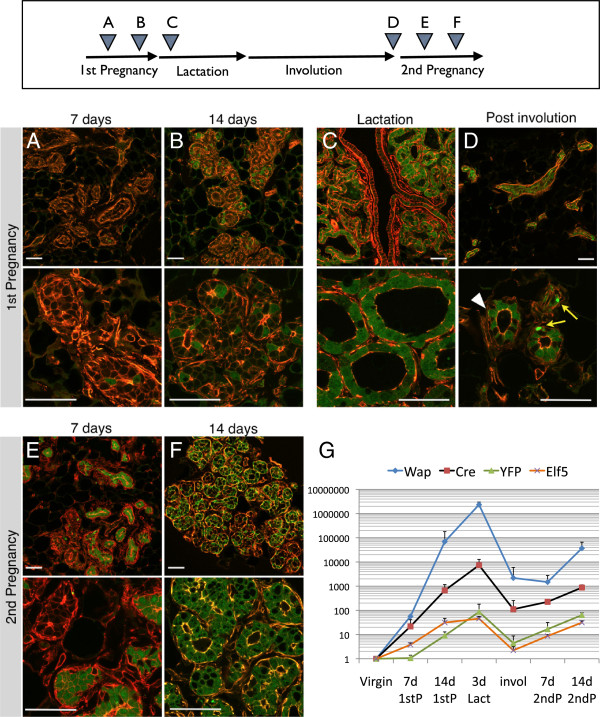
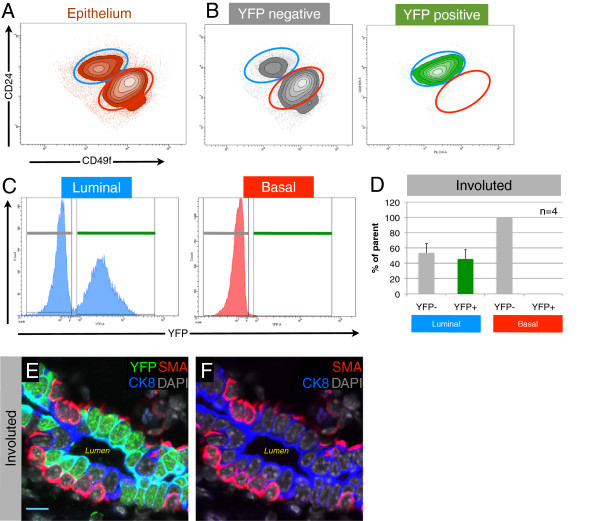
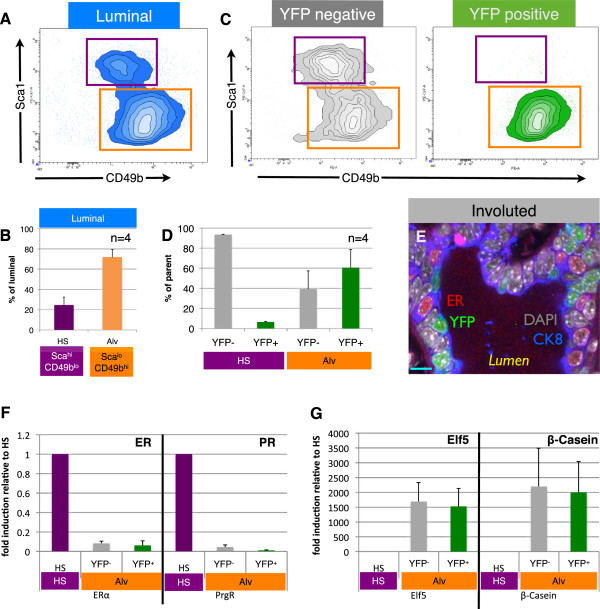
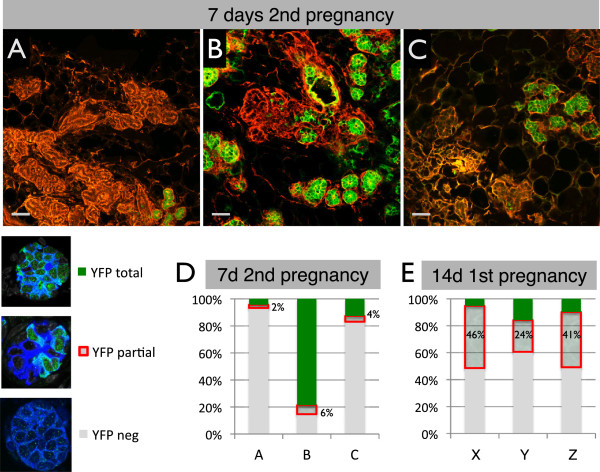
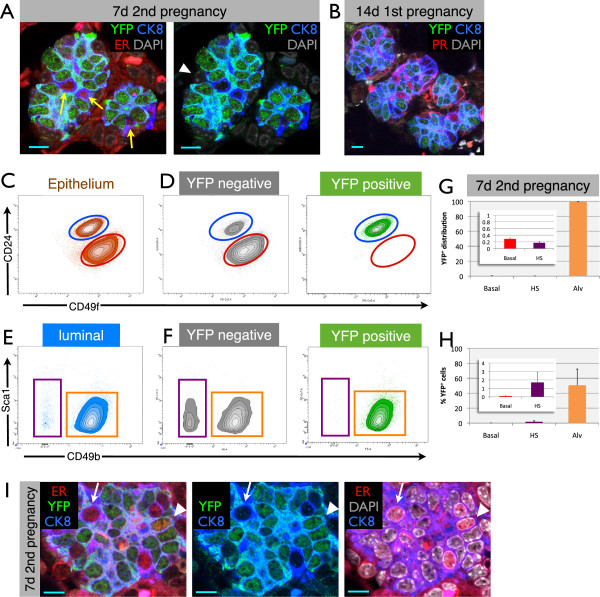
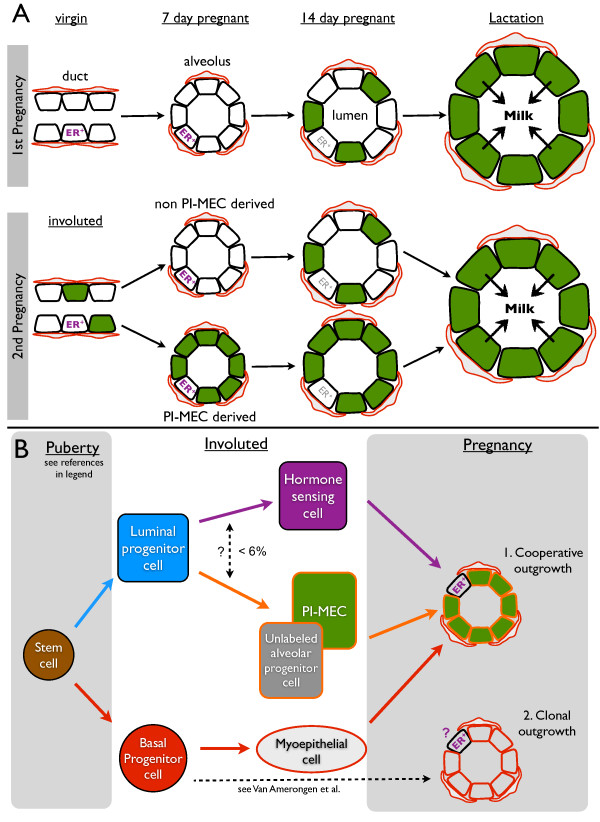
Results: After involution, PI-MECs are present exclusively in the luminal layer of mammary ducts. During pregnancy, PI-MECs contribute to the luminal layer but not the basal layer of alveolar lobules. Strikingly, whereas all luminal estrogen receptor (ER)-negative cells in an alveolus can be derived from PI-MECs, the alveolar ER-positive cells are unlabeled and reminiscent of Notch2-traced L cells. Notably, we observed a significant population of unlabeled alveolar progenitors that resemble PI-MECs based on transcriptional and histological analysis.
Conclusions: Our demonstration that PI-MECs are luminal cells underscores that not only basal cells display multi-lineage potential in transplantation assays. However, the lineage potential of PI-MECs in unperturbed mammary glands is remarkably restricted to luminal ER-negative cells of the secretory alveolar lineage. The identification of an unlabeled but functionally similar population of luminal alveolar progenitor cells raises the question of whether PI-MECs are a unique population or the result of stochastic labeling. Interestingly, even when all luminal ER-negative cells of an alveolus are PI-MEC-derived, the basal cells and hormone-sensing cells are derived from a different source, indicating that cooperative outgrowth of cells from different lineages is common in alveologenesis.
Figures
Similar articles
-
Parity-induced mammary epithelial cells are multipotent and express cell surface markers associated with stem cells.Dev Biol. 2007 Mar 1;303(1):29-44. doi: 10.1016/j.ydbio.2006.12.017. Epub 2006 Dec 13. Dev Biol. 2007. PMID: 17222404
-
Evidence for a multipotent mammary progenitor with pregnancy-specific activity.Breast Cancer Res. 2013;15(4):R65. doi: 10.1186/bcr3459. Breast Cancer Res. 2013. PMID: 23947835 Free PMC article.
-
Notch-induced mammary tumorigenesis does not involve the lobule-limited epithelial progenitor.Oncogene. 2012 Jan 5;31(1):60-7. doi: 10.1038/onc.2011.215. Epub 2011 Jun 13. Oncogene. 2012. PMID: 21666720 Free PMC article.
-
Does the Mouse Mammary Gland Arise from Unipotent or Multipotent Mammary Stem/Progenitor Cells?J Mammary Gland Biol Neoplasia. 2018 Jun;23(1-2):1-3. doi: 10.1007/s10911-018-9394-2. Epub 2018 Apr 11. J Mammary Gland Biol Neoplasia. 2018. PMID: 29644495 Review.
-
The myoepithelial cell: its role in normal mammary glands and breast cancer.Folia Morphol (Warsz). 2010 Feb;69(1):1-14. Folia Morphol (Warsz). 2010. PMID: 20235044 Review.
Cited by
-
Luminal epithelial cells within the mammary gland can produce basal cells upon oncogenic stress.Oncogene. 2016 Mar 17;35(11):1461-7. doi: 10.1038/onc.2015.206. Epub 2015 Jun 22. Oncogene. 2016. PMID: 26096929 Free PMC article.
-
Cellular Plasticity of Mammary Epithelial Cells Underlies Heterogeneity of Breast Cancer.Biomedicines. 2018 Nov 1;6(4):103. doi: 10.3390/biomedicines6040103. Biomedicines. 2018. PMID: 30388868 Free PMC article. Review.
-
Got Milk? Identifying and Characterizing Lactation Defects in Genetically-Engineered Mouse Models.J Mammary Gland Biol Neoplasia. 2020 Dec;25(4):255-272. doi: 10.1007/s10911-020-09467-y. Epub 2020 Nov 19. J Mammary Gland Biol Neoplasia. 2020. PMID: 33211270 Review.
-
Mammary lineage tracing: the coming of age.Cell Mol Life Sci. 2015 Apr;72(8):1577-83. doi: 10.1007/s00018-014-1817-7. Epub 2015 Jan 7. Cell Mol Life Sci. 2015. PMID: 25563489 Free PMC article. Review.
-
Lineage Tracing Methods to Study Mammary Epithelial Hierarchies In Vivo.Methods Mol Biol. 2022;2471:141-157. doi: 10.1007/978-1-0716-2193-6_7. Methods Mol Biol. 2022. PMID: 35175595
References
-
- Regan JL, Sourisseau T, Soady K, Kendrick H, McCarthy A, Tang C, Brennan K, Linardopoulos S, White DE, Smalley MJ. Aurora a kinase regulates mammary epithelial cell fate by determining mitotic spindle orientation in a notch-dependent manner. Cell Rep. 2013;4:110–123. doi: 10.1016/j.celrep.2013.05.044. - DOI - PubMed
-
- Kordon E, Smith G. An entire functional mammary gland may comprise the progeny from a single cell. Development. 1998;125:1921–1930. - PubMed
-
- Perou CM, Sørlie T, Eisen MB, van de Rijn M, Jeffrey SS, Rees CA, Pollack JR, Ross DT, Johnsen H, Akslen LA, Fluge O, Pergamenschikov A, Williams C, Zhu SX, Lønning PE, Børresen-Dale AL, Brown PO, Botstein D. Molecular portraits of human breast tumours. Nature. 2000;406:747–752. doi: 10.1038/35021093. - DOI - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
