

Pyroptotic neuronal cell death mediated by the AIM2 inflammasome
- PMID: 24398937
- PMCID: PMC3982080
- DOI: 10.1038/jcbfm.2013.236
Pyroptotic neuronal cell death mediated by the AIM2 inflammasome
Erratum in
-
Corrigendum.J Cereb Blood Flow Metab. 2016 Feb;36(2):456. doi: 10.1177/0271678X15620125. J Cereb Blood Flow Metab. 2016. PMID: 26814199 Free PMC article.
Abstract
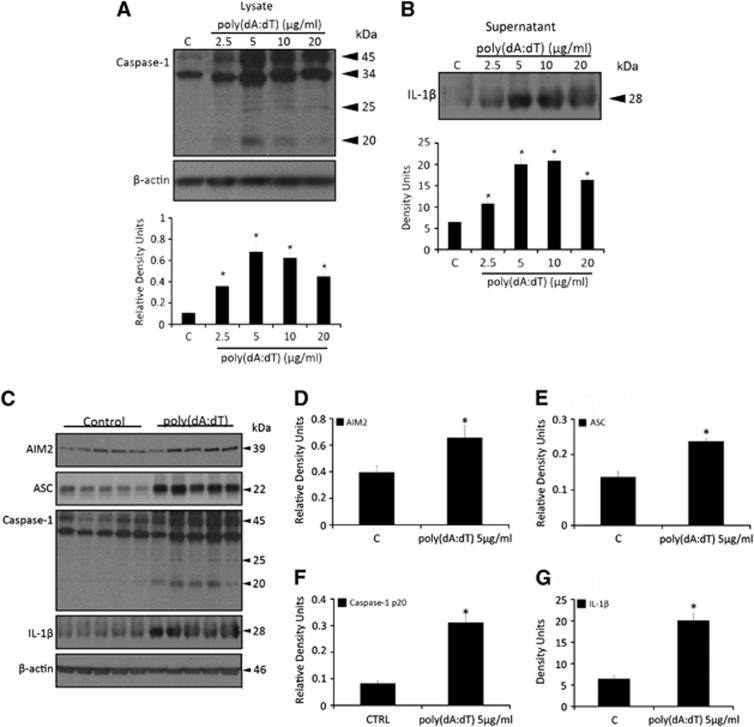
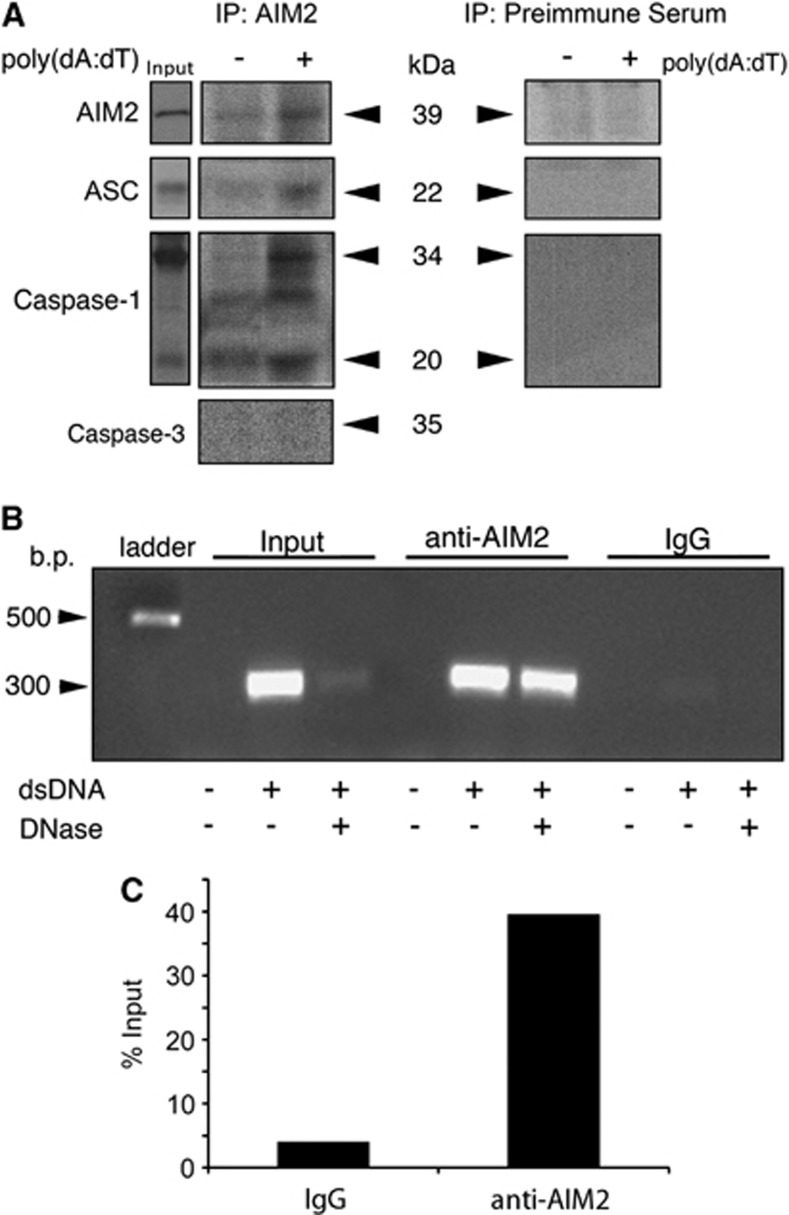
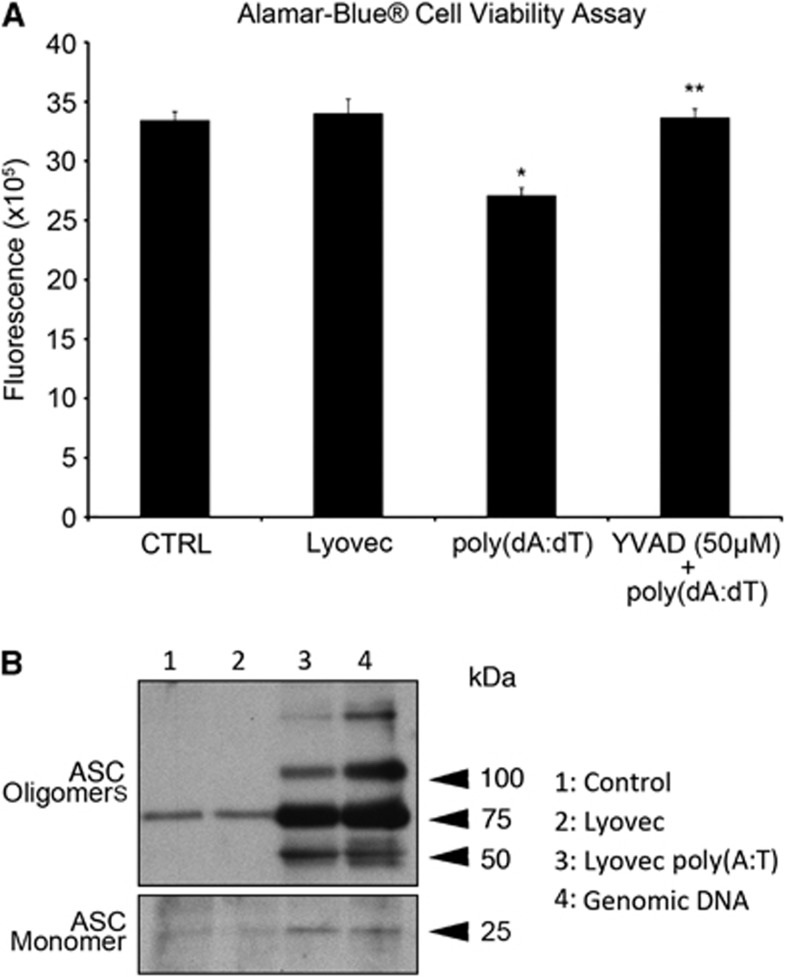
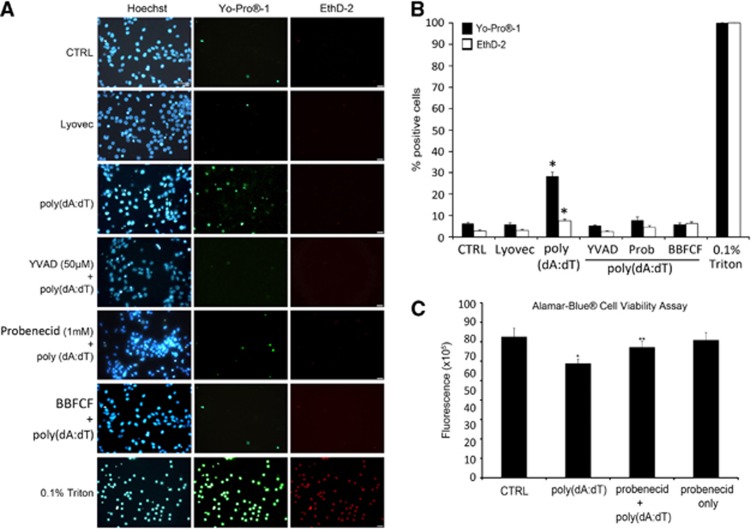
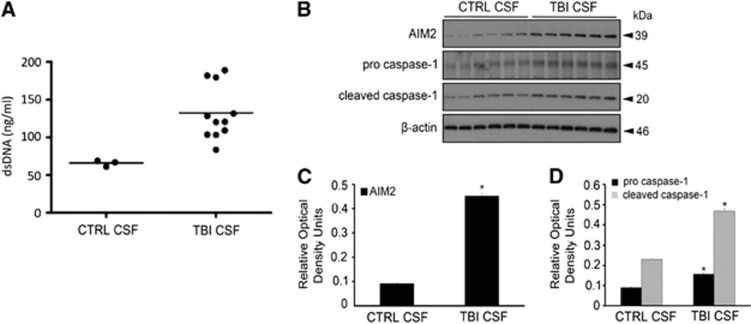
The central nervous system (CNS) is an active participant in the innate immune response to infection and injury. In these studies, we show embryonic cortical neurons express a functional, deoxyribonucleic acid (DNA)-responsive, absent in melanoma 2 (AIM2) inflammasome that activates caspase-1. Neurons undergo pyroptosis, a proinflammatory cell death mechanism characterized by the following: (a) oligomerization of apoptosis-associated speck-like protein containing a caspase recruitment domain (ASC); (b) caspase-1 dependency; (c) formation of discrete pores in the plasma membrane; and (d) release of the inflammatory cytokine interleukin-1β (IL-1β). Probenecid and Brilliant Blue FCF, inhibitors of the pannexin1 channel, prevent AIM2 inflammasome-mediated cell death, identifying pannexin1 as a cell death effector during pyroptosis and probenecid as a novel pyroptosis inhibitor. Furthermore, we show activation of the AIM2 inflammasome in neurons by cerebrospinal fluid (CSF) from traumatic brain injury (TBI) patients and oligomerization of ASC. These findings suggest neuronal pyroptosis is an important cell death mechanism during CNS infection and injury that may be attenuated by probenecid.
Figures

References
-
- Martinon F, Burns K, Tschopp J. The inflammasome: a molecular platform triggering activation of inflammatory caspases and processing of proIL-beta. Mol Cell. 2002;10:417–426. - PubMed
-
- Halle M, Gabrielsen A, Paulsson-Berne G, Gahm C, Agardh HE, Farnebo F, et al. Sustained inflammation due to nuclear factor-kappa B activation in irradiated human arteries. J Am Coll Cardiol. 2010;55:1227–1236. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
