

Recovery after brain injury: mechanisms and principles
- PMID: 24399951
- PMCID: PMC3870954
- DOI: 10.3389/fnhum.2013.00887
Recovery after brain injury: mechanisms and principles
Abstract
The past 20 years have represented an important period in the development of principles underlying neuroplasticity, especially as they apply to recovery from neurological injury. It is now generally accepted that acquired brain injuries, such as occur in stroke or trauma, initiate a cascade of regenerative events that last for at least several weeks, if not months. Many investigators have pointed out striking parallels between post-injury plasticity and the molecular and cellular events that take place during normal brain development. As evidence for the principles and mechanisms underlying post-injury neuroplasticity has been gleaned from both animal models and human populations, novel approaches to therapeutic intervention have been proposed. One important theme has persisted as the sophistication of clinicians and scientists in their knowledge of neuroplasticity mechanisms has grown: behavioral experience is the most potent modulator of brain plasticity. While there is substantial evidence for this principle in normal, healthy brains, the injured brain is particularly malleable. Based on the quantity and quality of motor experience, the brain can be reshaped after injury in either adaptive or maladaptive ways. This paper reviews selected studies that have demonstrated the neurophysiological and neuroanatomical changes that are triggered by motor experience, by injury, and the interaction of these processes. In addition, recent studies using new and elegant techniques are providing novel perspectives on the events that take place in the injured brain, providing a real-time window into post-injury plasticity. These new approaches are likely to accelerate the pace of basic research, and provide a wealth of opportunities to translate basic principles into therapeutic methodologies.
Keywords: axonal sprouting; motor cortex; motor learning; recovery; stroke; traumatic brain injury.
Figures
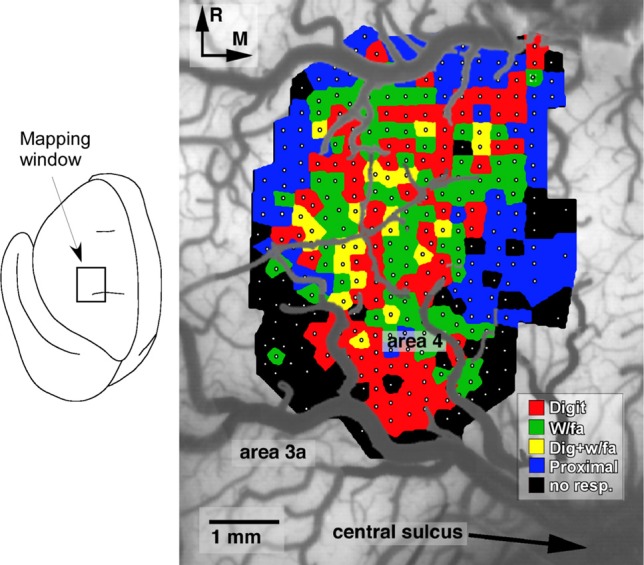
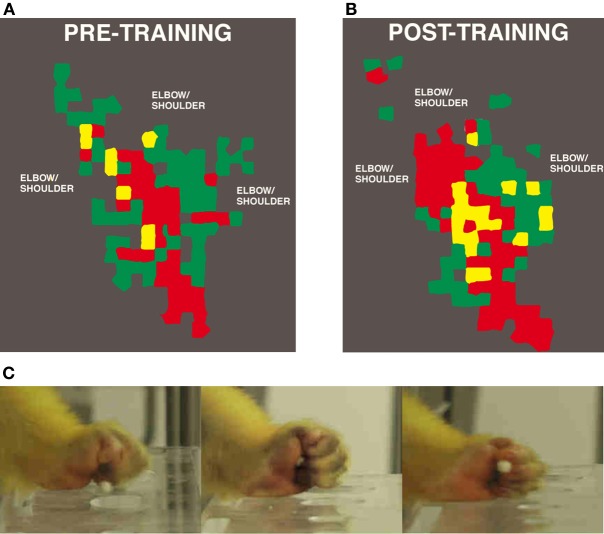
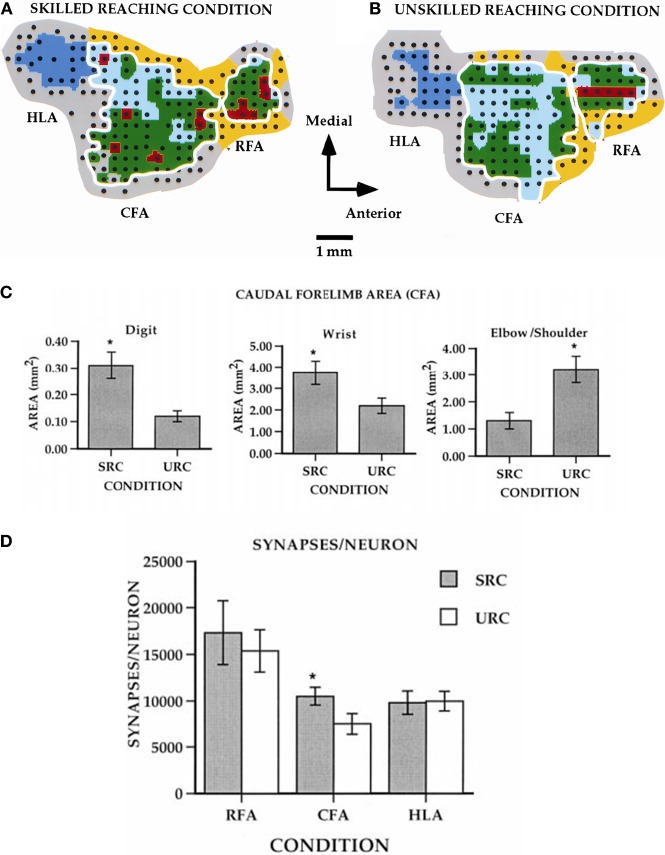
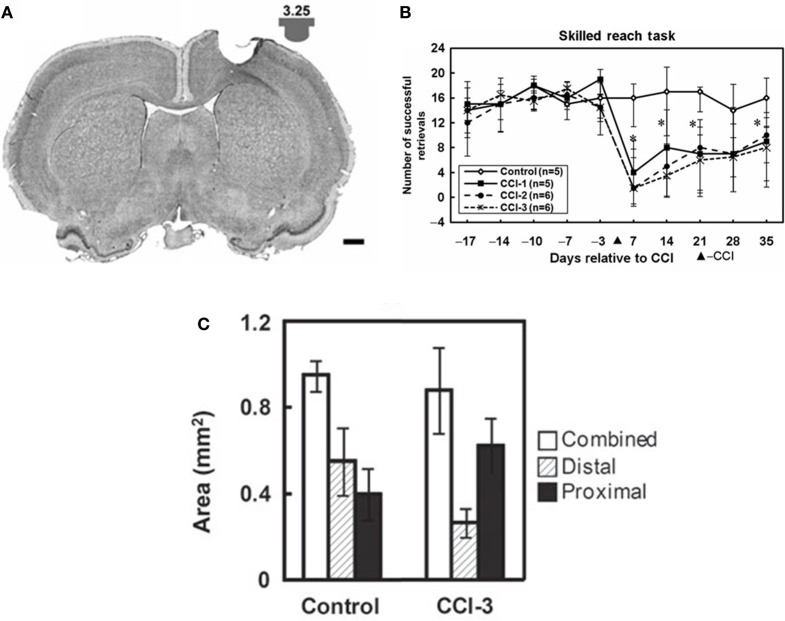
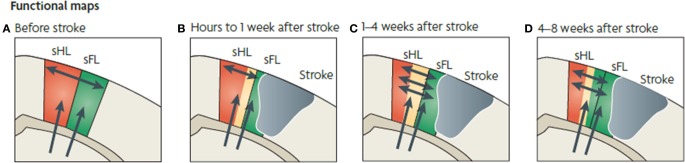
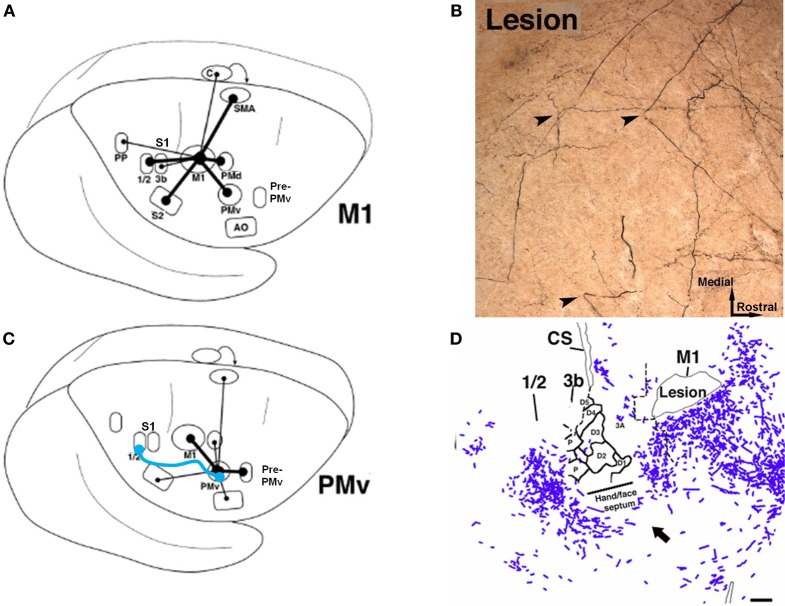
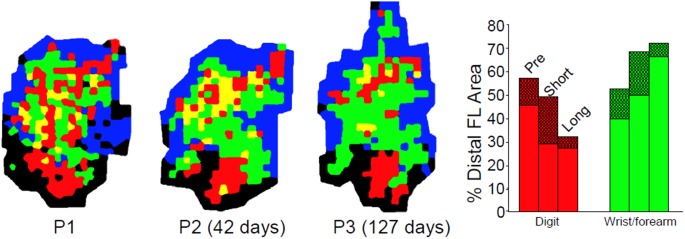
References
-
- Allred R. P., Maldonado M. A., Hsu J.e., Jones T. A. (2005). Training the “less-affected” forelimb after unilateral cortical infarcts interferes with functional recovery of the impaired forelimb in rats. Restor. Neurol. Neurosci. 23, 297–302 - PubMed
-
- Aroniadou V. A., Keller A. (1993). The patterns and synaptic properties of horizontal intracortical connections in the rat motor cortex. J. Neurophysiol. 70, 1553–1569 - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous