

Evolution of mammalian Opn5 as a specialized UV-absorbing pigment by a single amino acid mutation
- PMID: 24403072
- PMCID: PMC3924266
- DOI: 10.1074/jbc.M113.514075
Evolution of mammalian Opn5 as a specialized UV-absorbing pigment by a single amino acid mutation
Abstract
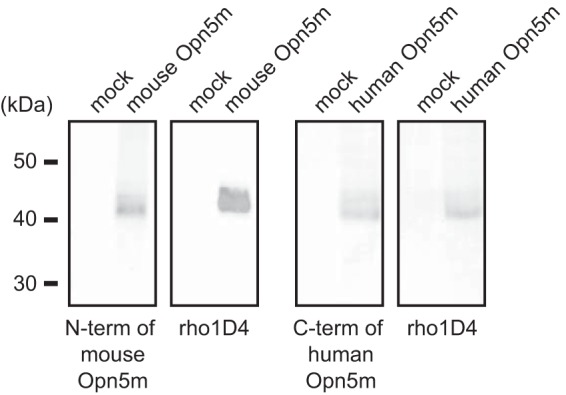
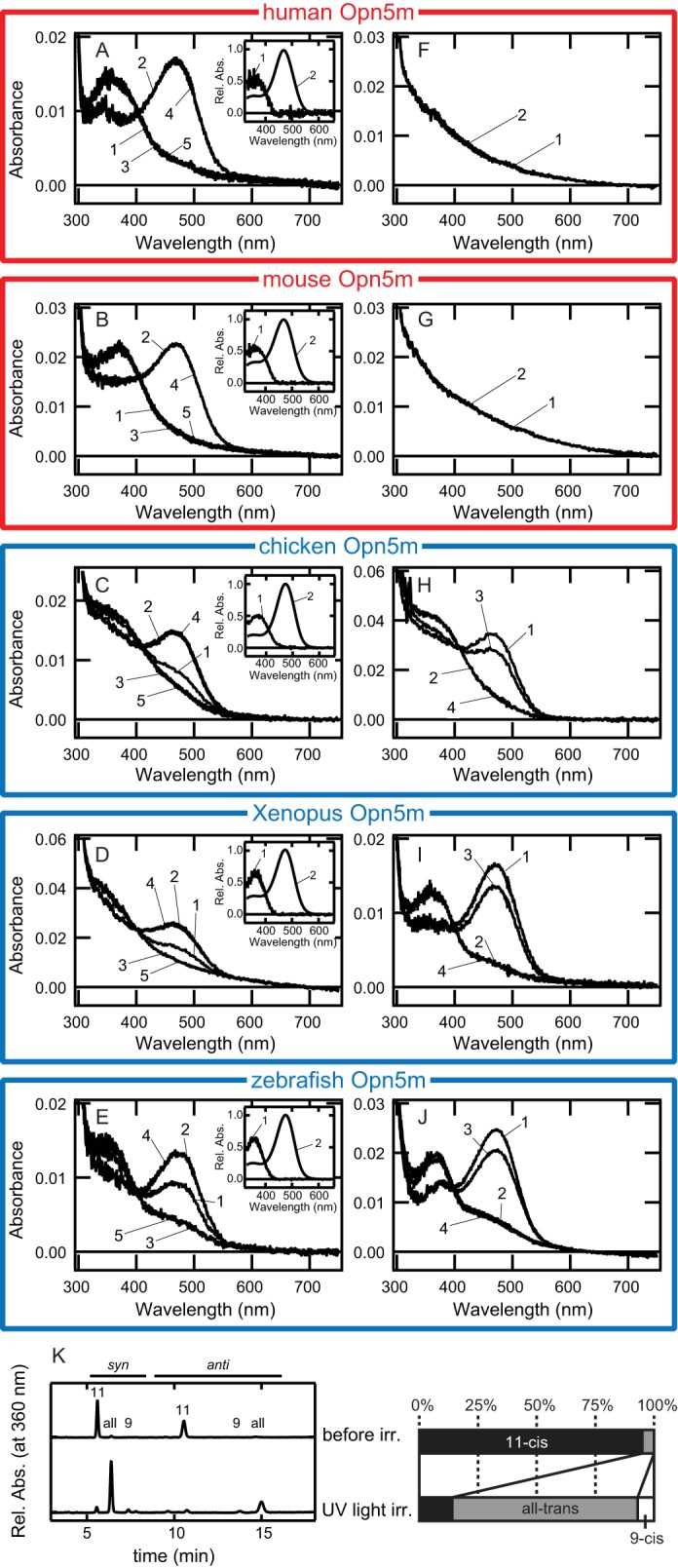
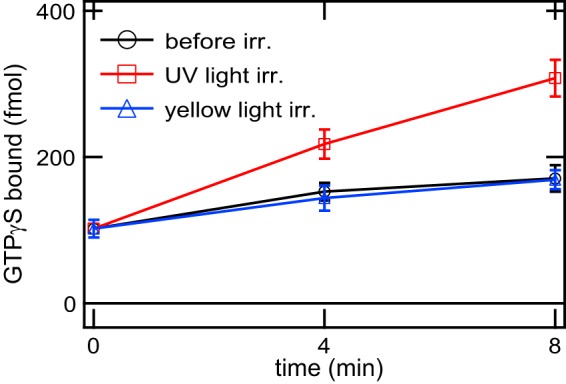
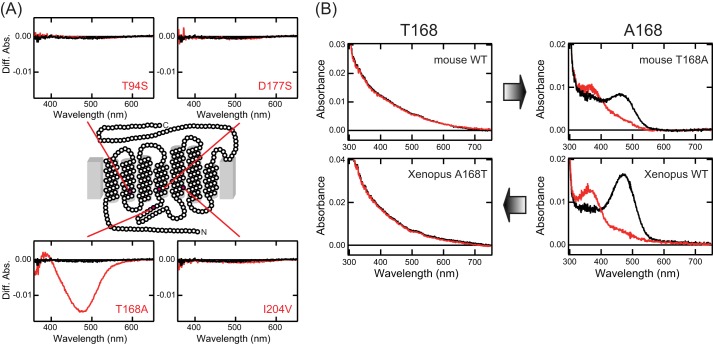
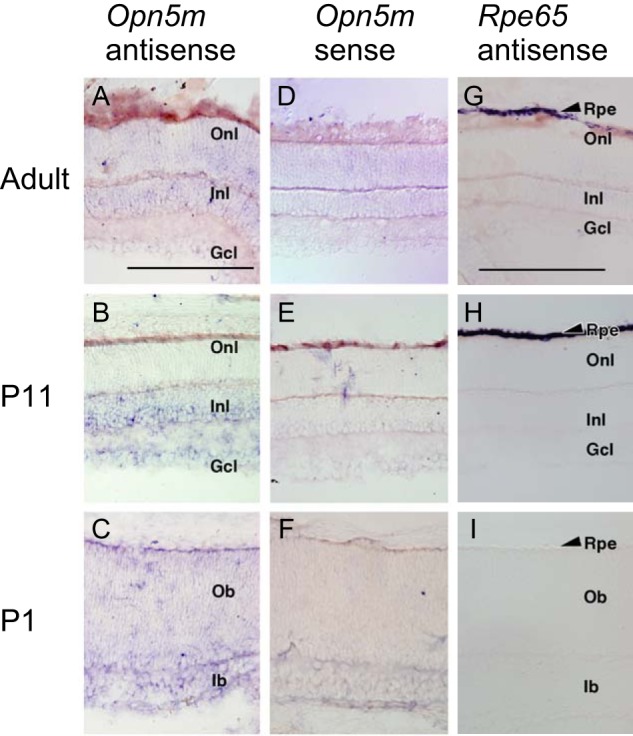
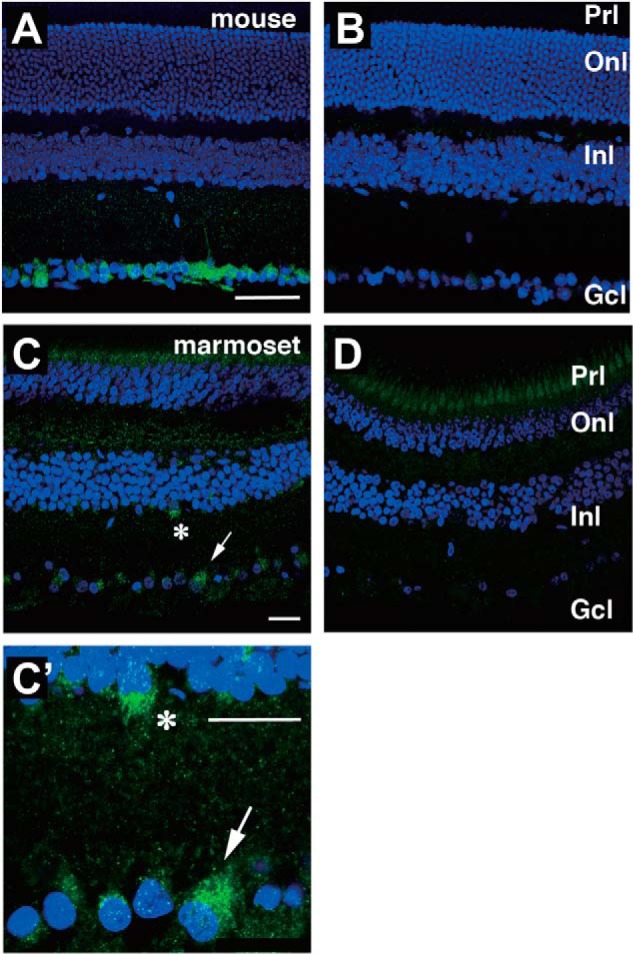
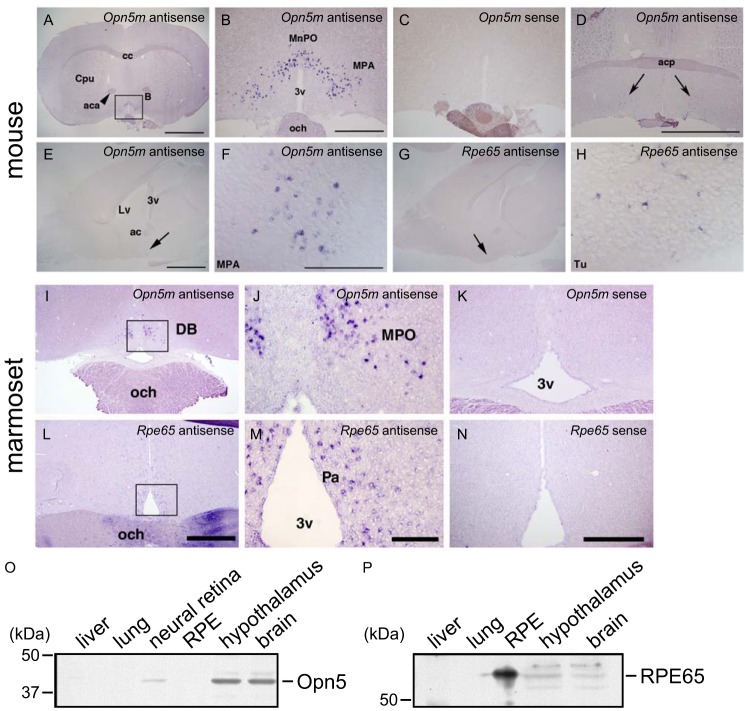
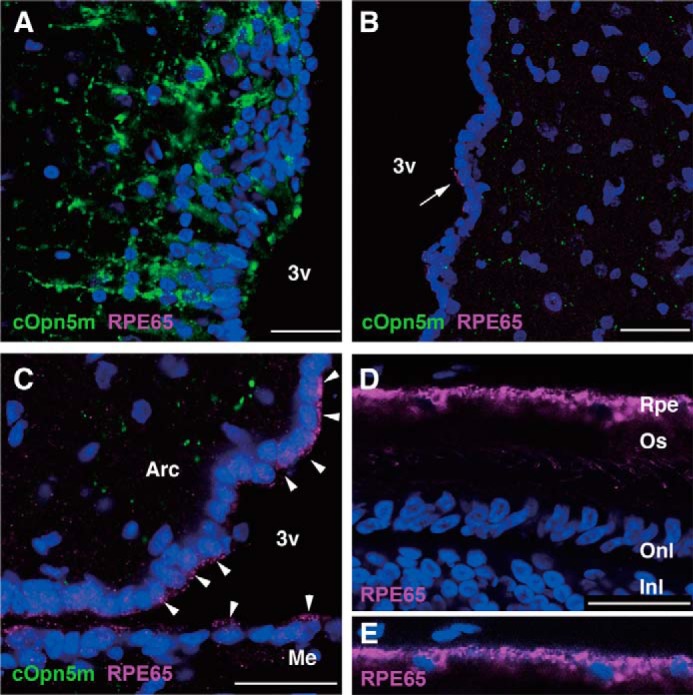
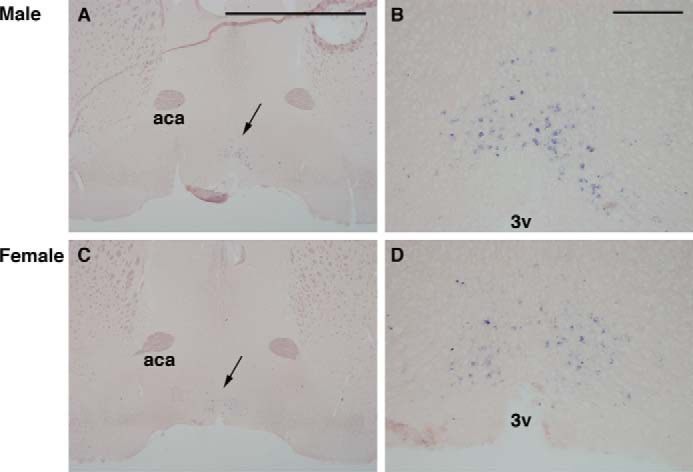
Opn5 is one of the recently identified opsin groups that is responsible for nonvisual photoreception in animals. We previously showed that a chicken homolog of mammalian Opn5 (Opn5m) is a Gi-coupled UV sensor having molecular properties typical of bistable pigments. Here we demonstrated that mammalian Opn5m evolved to be a more specialized photosensor by losing one of the characteristics of bistable pigments, direct binding of all-trans-retinal. We first confirmed that Opn5m proteins in zebrafish, Xenopus tropicalis, mouse, and human are also UV-sensitive pigments. Then we found that only mammalian Opn5m proteins lack the ability to directly bind all-trans-retinal. Mutational analysis showed that these characteristics were acquired by a single amino acid replacement at position 168. By comparing the expression patterns of Opn5m between mammals and chicken, we found that, like chicken Opn5m, mammalian Opn5m was localized in the ganglion cell layer and inner nuclear layer of the retina. However, the mouse and primate (common marmoset) opsins were distributed not in the posterior hypothalamus (including the region along the third ventricle) where chicken Opn5m is localized, but in the preoptic hypothalamus. Interestingly, RPE65, an essential enzyme for forming 11-cis-retinal in the visual cycle is expressed near the preoptic hypothalamus of the mouse and common marmoset brain but not near the region of the chicken brain where chicken Opn5m is expressed. Therefore, mammalian Opn5m may work exclusively as a short wavelength sensor in the brain as well as in the retina with the assistance of an 11-cis-retinal-supplying system.
Keywords: G Proteins; Molecular Evolution; Nonvisual Photoreception; Photoreceptors; Rhodopsin; Signal Transduction.
Figures
References
-
- Koyanagi M., Terakita A. (2008) Gq-coupled rhodopsin subfamily composed of invertebrate visual pigment and melanopsin. Photochem. Photobiol. 84, 1024–1030 - PubMed
-
- Tarttelin E. E., Bellingham J., Hankins M. W., Foster R. G., Lucas R. J. (2003) Neuropsin (Opn5). A novel opsin identified in mammalian neural tissue. FEBS Lett. 554, 410–416 - PubMed
-
- Tomonari S., Migita K., Takagi A., Noji S., Ohuchi H. (2008) Expression patterns of the opsin 5-related genes in the developing chicken retina. Dev. Dyn. 237, 1910–1922 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
