

Restraint stress alters nociceptin/orphanin FQ and CRF systems in the rat central amygdala: significance for anxiety-like behaviors
- PMID: 24403138
- PMCID: PMC3870926
- DOI: 10.1523/JNEUROSCI.2400-13.2014
Restraint stress alters nociceptin/orphanin FQ and CRF systems in the rat central amygdala: significance for anxiety-like behaviors
Abstract
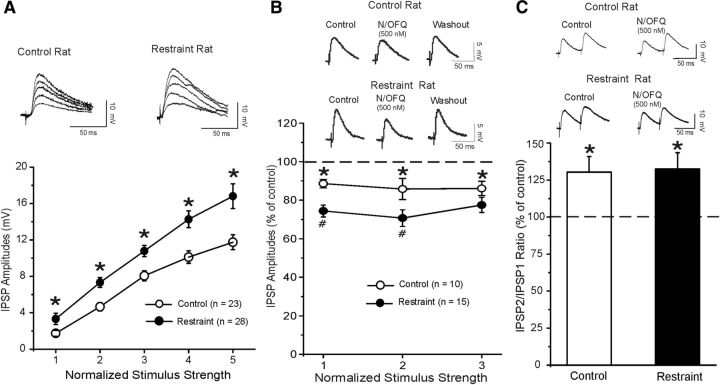
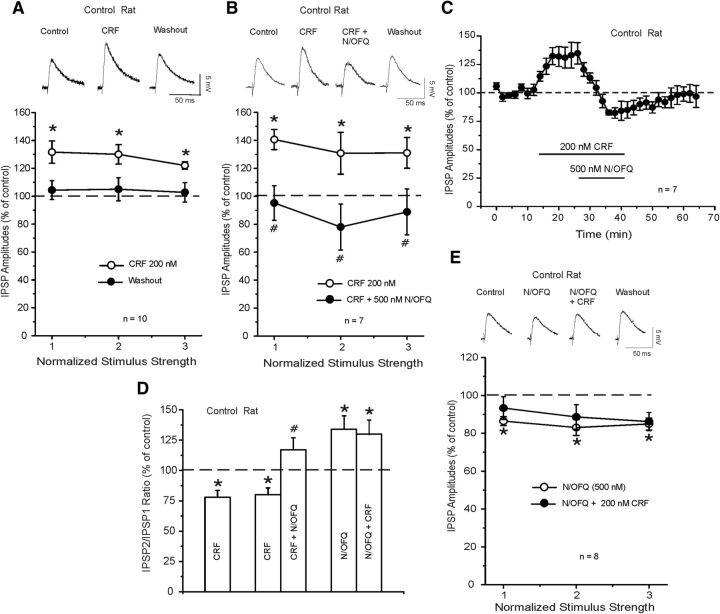
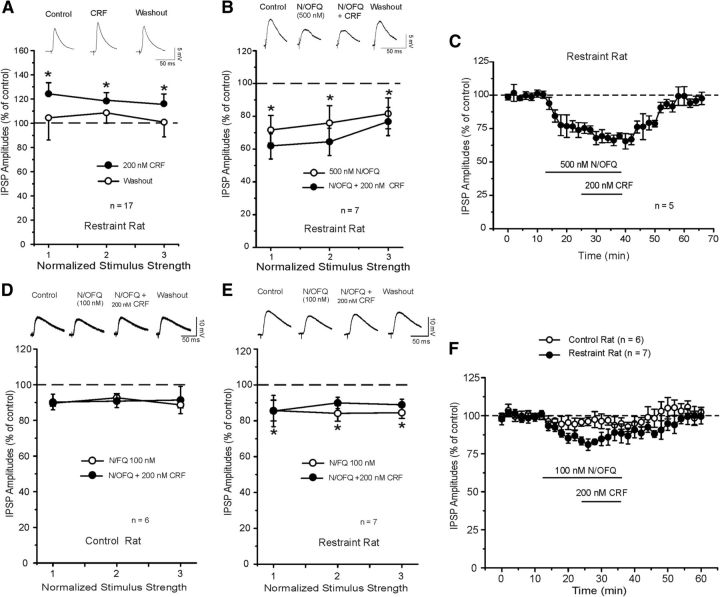
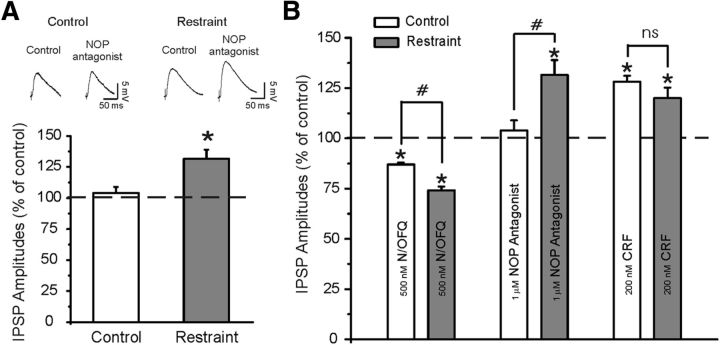
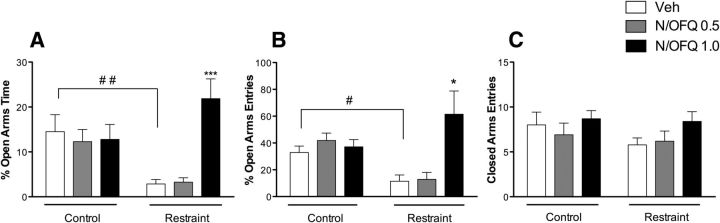
Corticotropin releasing factor (CRF) is the primary mediator of stress responses, and nociceptin/orphanin FQ (N/OFQ) plays an important role in the modulation of these stress responses. Thus, in this multidisciplinary study, we explored the relationship between the N/OFQ and the CRF systems in response to stress. Using in situ hybridization (ISH), we assessed the effect of body restraint stress on the gene expression of CRF and N/OFQ-related genes in various subdivisions of the amygdala, a critical brain structure involved in the modulation of stress response and anxiety-like behaviors. We found a selective upregulation of the NOP and downregulation of the CRF1 receptor transcripts in the CeA and in the BLA after body restraint. Thus, we performed intracellular electrophysiological recordings of GABAA-mediated IPSPs in the central nucleus of the amygdala (CeA) to explore functional interactions between CRF and N/OFQ systems in this brain region. Acute application of CRF significantly increased IPSPs in the CeA, and this enhancement was blocked by N/OFQ. Importantly, in stress-restraint rats, baseline CeA GABAergic responses were elevated and N/OFQ exerted a larger inhibition of IPSPs compared with unrestraint rats. The NOP antagonist [Nphe1]-nociceptin(1-13)NH2 increased the IPSP amplitudes in restraint rats but not in unrestraint rats, suggesting a functional recruitment of the N/OFQ system after acute stress. Finally, we evaluated the anxiety-like response in rats subjected to restraint stress and nonrestraint rats after N/OFQ microinjection into the CeA. Intra-CeA injections of N/OFQ significantly and selectively reduced anxiety-like behavior in restraint rats in the elevated plus maze. These combined results demonstrate that acute stress increases N/OFQ systems in the CeA and that N/OFQ has antistress properties.
Figures
References
-
- Aujla H, Cannarsa R, Romualdi P, Ciccocioppo R, Martin-Fardon R, Weiss F. Modification of anxiety-like behaviors by nociceptin/orphanin FQ (N/OFQ) and time-dependent changes in N/OFQ-NOP gene expression following ethanol withdrawal. Addict Biol. 2013;18:467–479. doi: 10.1111/j.1369-1600.2012.00466.x. - DOI - PMC - PubMed
-
- Ciccocioppo R, Economidou D, Fedeli A, Angeletti S, Weiss F, Heilig M, Massi M. Attenuation of ethanol self-administration and of conditioned reinstatement of alcohol-seeking behaviour by the antiopioid peptide nociceptin/orphanin FQ in alcohol-preferring rats. Psychopharmacology (Berl) 2004;172:170–178. doi: 10.1007/s00213-003-1645-1. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- U01 AA013498/AA/NIAAA NIH HHS/United States
- AA015566/AA/NIAAA NIH HHS/United States
- AA013498/AA/NIAAA NIH HHS/United States
- AA021491/AA/NIAAA NIH HHS/United States
- R01 AA017447/AA/NIAAA NIH HHS/United States
- P60 AA006420/AA/NIAAA NIH HHS/United States
- R01 AA015566/AA/NIAAA NIH HHS/United States
- R01 AA021491/AA/NIAAA NIH HHS/United States
- AA017447/AA/NIAAA NIH HHS/United States
- P50 AA006420/AA/NIAAA NIH HHS/United States
- AA06420/AA/NIAAA NIH HHS/United States
- AA016985/AA/NIAAA NIH HHS/United States
- R01 AA016985/AA/NIAAA NIH HHS/United States
- R01 AA014351/AA/NIAAA NIH HHS/United States
- AA014351/AA/NIAAA NIH HHS/United States
- R37 AA017447/AA/NIAAA NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical