

Insights into the structure and assembly of the bacteriophage 29 double-stranded DNA packaging motor
- PMID: 24403593
- PMCID: PMC3993773
- DOI: 10.1128/JVI.03203-13
Insights into the structure and assembly of the bacteriophage 29 double-stranded DNA packaging motor
Abstract
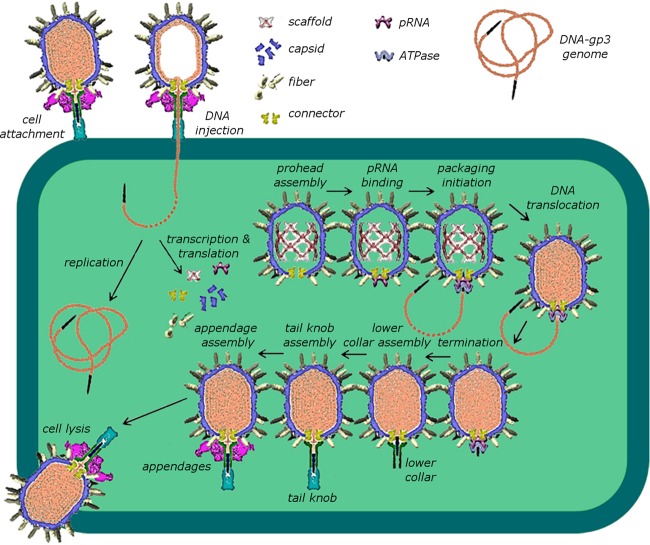
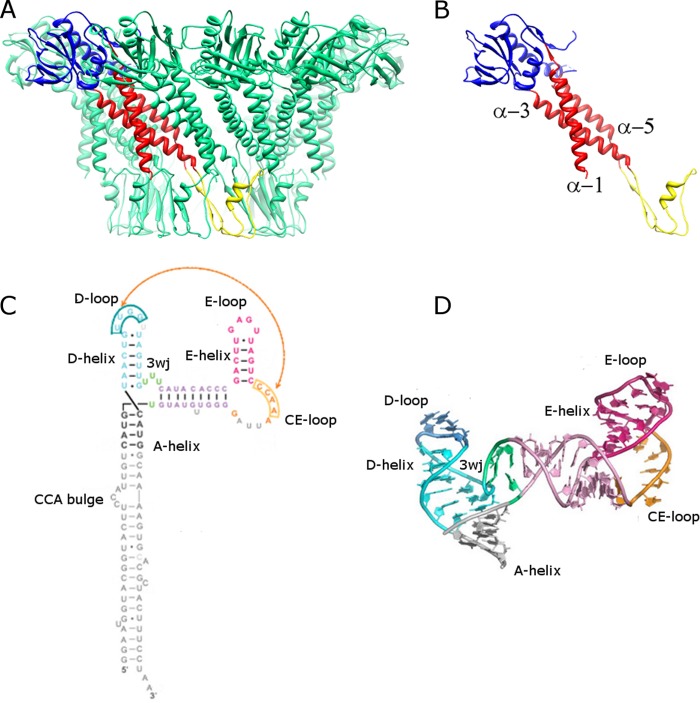
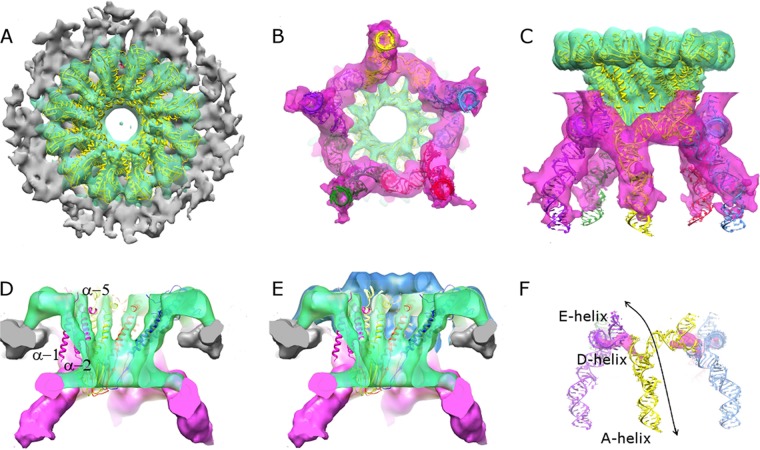
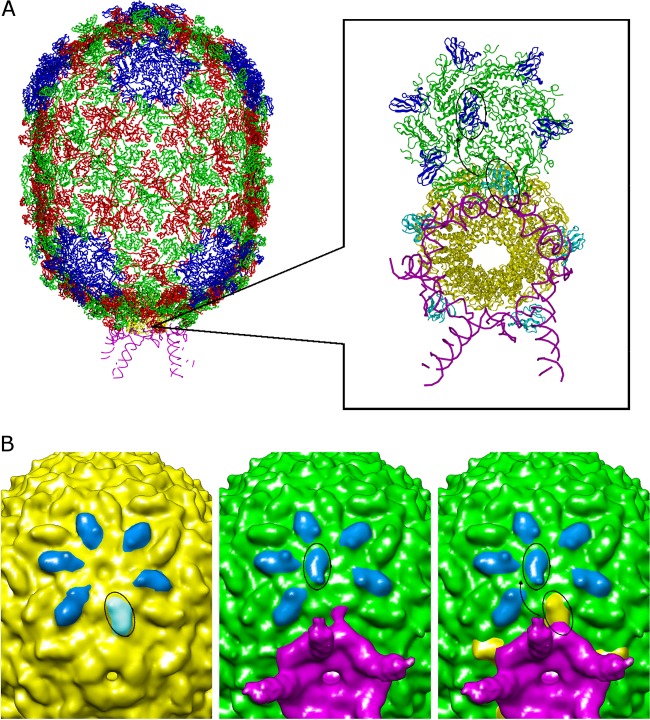
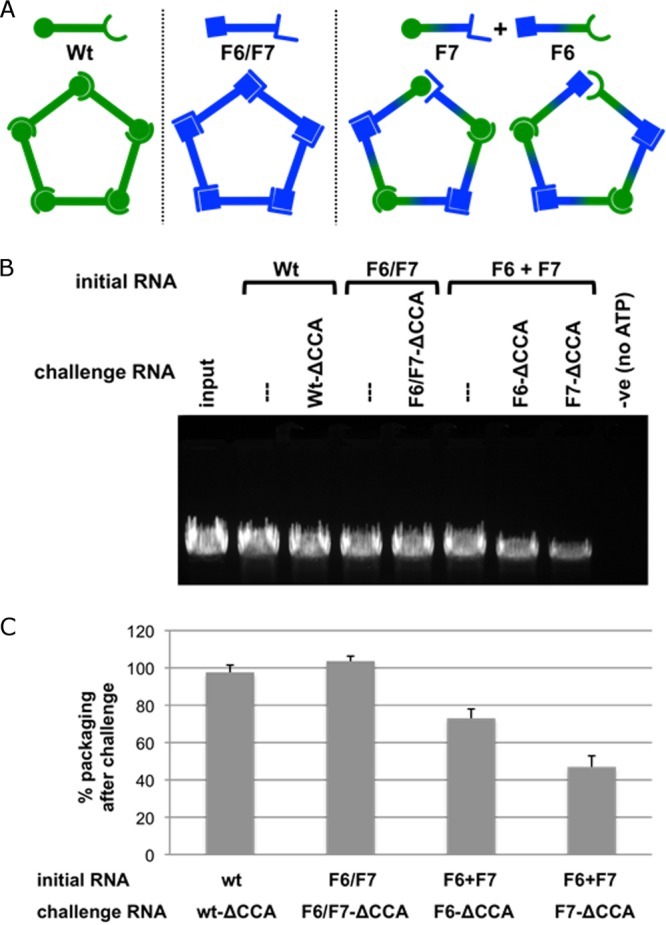
The tailed double-stranded DNA (dsDNA) bacteriophage 29 packages its 19.3-kbp genome into a preassembled procapsid structure by using a transiently assembled phage-encoded molecular motor. This process is remarkable considering that compaction of DNA to near-crystalline densities within the confined space of the capsid requires that the packaging motor work against significant entropic, enthalpic, and DNA-bending energies. The motor consists of three phage-encoded components: the dodecameric connector protein gp10, an oligomeric RNA molecule known as the prohead RNA (pRNA), and the homomeric ring ATPase gp16. Although atomic resolution structures of the connector and different pRNA subdomains have been determined, the mechanism of self-assembly and the resulting stoichiometry of the various motor components on the phage capsid have been the subject of considerable controversy. Here a subnanometer asymmetric cryoelectron microscopy (cryo-EM) reconstruction of a connector-pRNA complex at a unique vertex of the procapsid conclusively demonstrates the pentameric symmetry of the pRNA and illuminates the relative arrangement of the connector and the pRNA. Additionally, a combination of biochemical and cryo-EM analyses of motor assembly intermediates suggests a sequence of molecular events that constitute the pathway by which the motor assembles on the head, thereby reconciling conflicting data regarding pRNA assembly and stoichiometry. Taken together, these data provide new insight into the assembly, structure, and mechanism of a complex molecular machine.
Importance: Viruses consist of a protein shell, or capsid, that protects and surrounds their genetic material. Thus, genome encapsidation is a fundamental and essential step in the life cycle of any virus. In dsDNA viruses, powerful molecular motors essentially pump the viral DNA into a preformed protein shell. This article describes how a viral dsDNA packaging motor self-assembles on the viral capsid and provides insight into its mechanism of action.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
