

The spindle and kinetochore-associated (Ska) complex enhances binding of the anaphase-promoting complex/cyclosome (APC/C) to chromosomes and promotes mitotic exit
- PMID: 24403607
- PMCID: PMC3937086
- DOI: 10.1091/mbc.E13-07-0421
The spindle and kinetochore-associated (Ska) complex enhances binding of the anaphase-promoting complex/cyclosome (APC/C) to chromosomes and promotes mitotic exit
Abstract
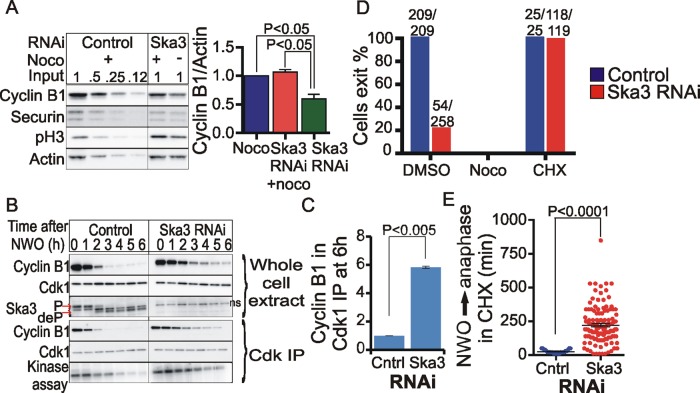
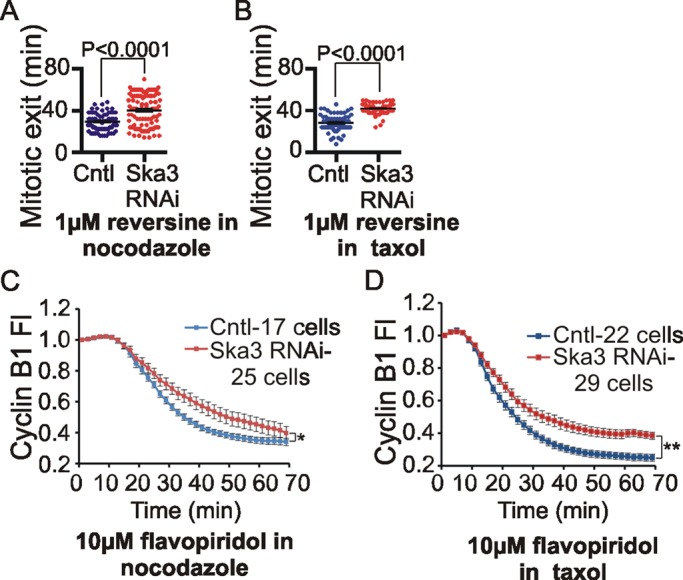
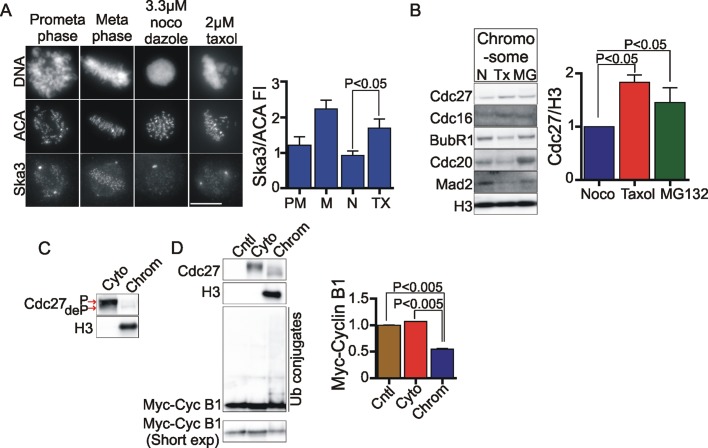
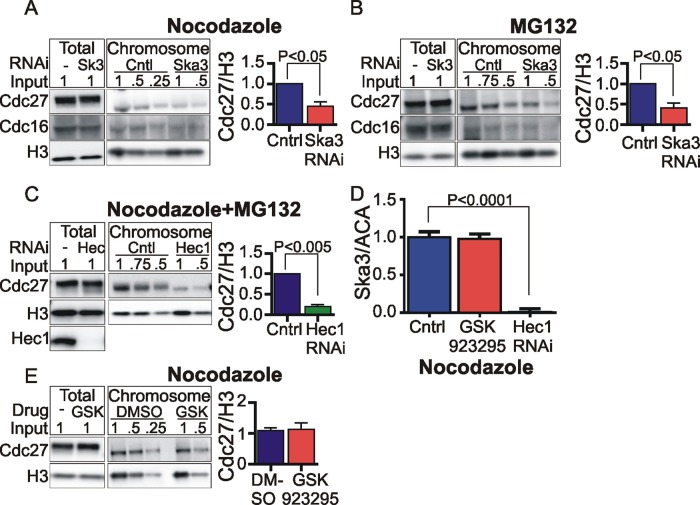
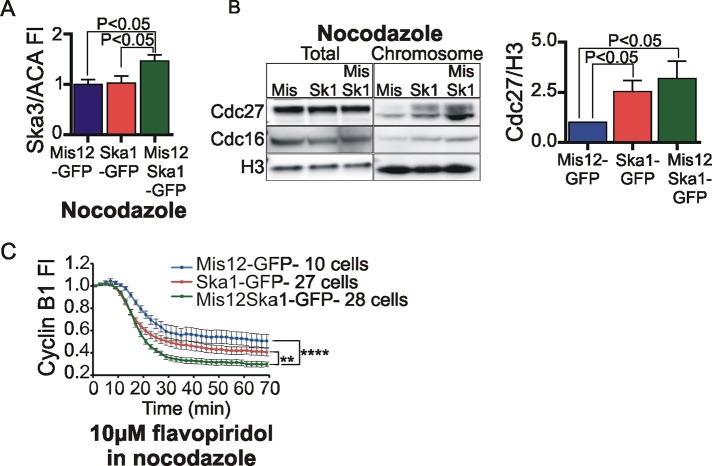
The spindle and kinetochore-associated (Ska) protein complex is a heterotrimeric complex required for timely anaphase onset. The major phenotypes seen after small interfering RNA-mediated depletion of Ska are transient alignment defects followed by metaphase arrest that ultimately results in cohesion fatigue. We find that cells depleted of Ska3 arrest at metaphase with only partial degradation of cyclin B1 and securin. In cells arrested with microtubule drugs, Ska3-depleted cells exhibit slower mitotic exit when the spindle checkpoint is silenced by inhibition of the checkpoint kinase, Mps1, or when cells are forced to exit mitosis downstream of checkpoint silencing by inactivation of Cdk1. These results suggest that in addition to a role in fostering kinetochore-microtubule attachment and chromosome alignment, the Ska complex has functions in promoting anaphase onset. We find that both Ska3 and microtubules promote chromosome association of the anaphase-promoting complex/cyclosome (APC/C). Chromosome-bound APC/C shows significantly stronger ubiquitylation activity than cytoplasmic APC/C. Forced localization of Ska complex to kinetochores, independent of microtubules, results in enhanced accumulation of APC/C on chromosomes and accelerated cyclin B1 degradation during induced mitotic exit. We propose that a Ska-microtubule-kinetochore association promotes APC/C localization to chromosomes, thereby enhancing anaphase onset and mitotic exit.
Figures

References
-
- Acquaviva C, Herzog F, Kraft C, Pines J. The anaphase promoting complex/cyclosome is recruited to centromeres by the spindle assembly checkpoint. Nat Cell Biol. 2004;6:892–898. - PubMed
-
- Clute P, Pines J. Temporal and spatial control of cyclin B1 destruction in metaphase. Nat Cell Biol. 1999;1:82–87. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
